Frontiers in Psychology
On September 28, 2021 by adminDas Corpus callosum (CC) ist die primäre Kommissur, die die beiden Gehirnhälften verbindet. Daher ist der Begriff „Kommissurotomie“ zur Beschreibung des so genannten „gespaltenen Gehirns“ zwar nicht falsch und für die ursprünglichen chirurgischen Eingriffe am Menschen, bei denen alle Kommissuren durchtrennt wurden, absolut korrekt (Gazzaniga et al., 1967), spätere Operationen durchtrennten nur das CC und brachten so den Begriff „Callosotomie“ in Gebrauch, der in diesen Fällen der angemessenere Begriff bleibt (z.B. Franz et al., 1996, 2000).
Aus evolutionärer Sicht ist das CC eine neuere Entwicklung, die nur bei plazentalen Säugetieren beobachtet wurde (Katz et al., 1983; Mihrshahi, 2006). Man geht davon aus, dass die Funktion des ZNS zumindest teilweise von der vorderen Kommissur (AC) übernommen wurde, wie sie bei nicht plazentaren Säugetieren wie den Beuteltieren beobachtet wurde (Heath und Jones, 1971; Ashwell et al., 1996). Obwohl einige Restbahnen der AC im menschlichen Gehirn noch vorhanden sind, sind ihre Konnektivität und Struktur in der aktuellen Literatur noch nicht ausreichend untersucht, und die spärlichen Ergebnisse sind eher uneinheitlich (Lasco et al., 2002; Patel et al., 2010; Choi et al., 2011). So sind beispielsweise die Ergebnisse der Magnetresonanztomographie (MRT) uneinheitlich, was die Frage angeht, ob das AC bei Frauen und Männern ähnlich groß ist (Lasco et al., 2002; Patel et al., 2010). In einer relativ aktuellen Studie, in der die In-vivo-Technik der Diffusions-Tensor-Bildgebung (DTI) eingesetzt wurde, die sich die Diffusion von Wassermolekülen in Axonen zunutze macht, untersuchten Patel et al. (2010) acht Probanden und isolierten bei fünf Probanden erfolgreich Fasern des AC. Zu den Ergebnissen gehörte, dass bei einem Probanden ein großer Anteil der Fasern des hinteren Schenkels des AC bilateral in die parietalen Regionen wanderte, wo die meisten von ihnen endeten, während ein kleineres Bündel im präzentralen Gyrus endete und ein weiteres Bündel in den Temporallappen eintrat. Solche individuellen Unterschiede wurden bei allen Teilnehmern festgestellt, doch die Mehrheit der Probanden zeigte bilaterale Projektionen in den orbitofrontalen, parietalen und temporalen Kortex, in die der CC nicht eindeutig projiziert. Zusammengenommen deuten die Ergebnisse darauf hin, dass das AC sowohl primäre als auch rudimentäre Projektionen in neokortikale Bereiche hat, in die das CC nicht projiziert, wie auch andere vorgeschlagen haben (Di Virgilio et al., 1999; Patel et al., 2010).
Unserer Ansicht nach stützt sich das AAA in hohem Maße auf neokortikale Netzwerke und Konnektivität mit subkortikalen Strukturen wie den Basalganglien und dem Thalamus (neben anderen, die in unserem Vorschlag nicht speziell diskutiert werden) (Franz, 2012). Daher könnten alle evolutionären Entwicklungen, die den AC betreffen, auch eine gewisse Rolle im AAA-Modell spielen. Wir haben jedoch nur wenig Literatur gefunden, die sich speziell mit dieser möglichen Verbindung befasst.
Die Rolle des AC wurde bei Aufgaben untersucht, die wahrscheinlich Formen der Aufmerksamkeit erfordern, wie z. B. solche, die visuelle Aufmerksamkeit beinhalten. Es ist zum Beispiel bekannt, dass die Populationen des AC im Zusammenhang mit Aufgaben untersucht wurden, die visuelle Integration erfordern (Gazzaniga et al., 1962, 1965; Sperry, 1968; Levy et al., 1972). In jüngerer Zeit hat Corballis (1995) versucht, die visuelle Integration und die Aufmerksamkeit zu untersuchen, um frühere Diskrepanzen in der Akallusalforschung zu erklären. Corballis schlug ein zweigeteiltes visuelles Aufmerksamkeitssystem vor; ein Teil (der sich auf die Form bezieht) ist automatisch und in jeder Hemisphäre lokalisiert, der andere (der sich auf die Bewegung und den Ort bezieht) ist freiwillig und subkortikal. Das AAA-Modell kann auf dieser Beobachtung aufbauen und die in der Literatur beobachteten Diskrepanzen beheben. Nach dem von uns vorgeschlagenen Modell gibt es über mehrere Aufmerksamkeitsebenen hinweg andere Wege als den CC, die den Informationstransfer zwischen den Hemisphären vermitteln könnten. Eine Möglichkeit betrifft die Schaltkreise der Basalganglien, von denen wir annehmen, dass sie bei der Vermittlung der Aufmerksamkeitsteilung zwischen den Hemisphären von entscheidender Bedeutung sind (Franz, 2012). Durch die Durchtrennung des CC wird die direkte Übertragung der Aufmerksamkeit über kortikale Bahnen weitgehend unterbunden (es sei denn, das AC spielt eine Ersatzrolle). Die aufmerksamkeitsbasierte Integration visueller Informationen könnte jedoch immer noch über subkortikale Bahnen und/oder den AC vermittelt werden. Auf diese Weise könnte das von Corballis (1995) beobachtete zweiteilige Aufmerksamkeitssystem aufgrund der aufmerksamkeitsbasierten Vermittlung der visuellen Integration über das AC (und/oder die Basalganglien und die beteiligten Schaltkreise: Franz, 2012) vorhanden sein.
Studien, die sich mit dem Sehen und der Aufmerksamkeit befassen, müssen nicht völlig getrennt von dem System betrachtet werden, das die motorische Handlung reguliert. Nach unseren eigenen Erkenntnissen (Franz, 2004; Franz und Packman, 2004) sind verschiedene Formen der Aufmerksamkeit (visuell und intern-nicht-visuell) eng mit motorischen Handlungen verknüpft, und es liegt nahe, dass ein möglicher Schlüssel zum Verständnis einer möglichen Aufmerksamkeitsrolle des AC in der Verbindung zwischen Handlung, Sehen und Aufmerksamkeit liegt. In einer früheren Studie ließ Franz (2004) normale Teilnehmer bimanuell Kreise zeichnen, wobei das visuelle Feedback oder die interne Aufmerksamkeit auf jede Hand unterschiedlich manipuliert wurde. Die Größe der Kreise, die von einer Hand gezeichnet wurden, die eine Form von interner Aufmerksamkeit erhielt, wurde in ähnlicher Weise größer wie bei visuellem Feedback für diese Hand (im Vergleich zu keiner Aufmerksamkeit/keinem visuellen Feedback). Unser AAA-Modell impliziert, dass die Aufmerksamkeit eine Schlüsselkomponente bei der Regulierung des motorischen Outputs ist (sowie bei der Auswahl der geplanten motorischen Aktion), und soweit die vorliegenden Beweise reichen, können wir nicht ausschließen, dass die AC eine Rolle bei dieser Funktion spielt. Ein direkter Test der Rolle des AC bei kognitiven Aufgaben ist jedoch nicht trivial.
Die Prüfung, ob das AC an kognitiven Funktionen beteiligt ist, ist in der neuropsychologischen Forschung nahezu unmöglich, da Teilnehmer mit lokalisiertem AC nicht leicht zu finden sind (und uns keine bekannt sind). Es wäre daher viel intuitiver, neurologische Unterschiede im normalen AC zu bewerten und neurologische Daten mit Verhaltensdaten zu korrelieren. Die vorgestellte Traktographie-Methode (Aufklärung des AC) lässt sich relativ einfach auf große Stichproben der Normalbevölkerung anwenden und kann idealerweise mit der Leistung (basierend auf Aufgaben, die während separater Testsitzungen aus der DTI-Sammlung durchgeführt wurden) derselben Personen anhand von speziell entwickelten kognitiven Aufgaben kombiniert werden. Wir zeigen hier, dass DTI an einer kleinen Stichprobe von Personen aus der neurologisch normalen Bevölkerung durchgeführt werden kann und brauchbare Ergebnisse liefert. Der allgemeine Vorschlag wäre, ähnliche Methoden an einer größeren Stichprobe durchzuführen und (wie oben vorgeschlagen) die Ergebnisse (z. B. eines Proxy-Maßes für die Dichte des AC) mit Variablen bei einer kognitiven Aufgabe zu korrelieren.
In Abbildung 1 zeigen wir die Ergebnisse des DTI-Trackings an Daten von 10 weiblichen Teilnehmern, die aus einem öffentlichen DTI-Datensatz der Johns Hopkins University stammen (siehe Danksagungen für die vollständige Quelle). Das Durchschnittsalter betrug 22 Jahre (SD = 2,5). Die Bilder wurden mit der FSL Diffusion Toolbox (Smith et al., 2004; Woolrich et al., 2009; Jenkinson et al., 2012) verarbeitet, und die Verfahren entsprachen denen, die in früheren Studien zu anderen Faserbahnen (FDT; Behrens et al., 2003a,b, 2007; Johansen-Berg et al., 2004) verwendet wurden. Zunächst wurden die Bilder für jeden Teilnehmer zu 4d-Volumina konkateniert und die Schädel mit dem FSL Brain Extraction Tool (BET; Smith, 2002; Jenkinson et al., 2005) extrahiert. Die extrahierten Bilder wurden um geringfügige Kopfbewegungen und Wirbelströme korrigiert, die durch Änderungen der Feldrichtung erzeugt wurden. Die Diffusionstensoren wurden angepasst, um Bilder zu erzeugen, die die primären Diffusionsrichtungen und die FA-Werte an jedem Voxel anzeigen, die für Farbkarten während der Maskierung verwendet werden. Eine Bayes’sche Schätzung von Diffusionsparametern, die unter Verwendung von Sampling-Techniken mit sich kreuzenden Fasern (BEDPOSTX; „X“ steht für sich kreuzende Fasern) erhalten wurden, wurde durchgeführt, um eine Verteilung der Diffusionsgröße und -richtung zu erzeugen, die zwei sich kreuzende Fasern berücksichtigt, um später eine Traktographie über Diffusionsvoxel durchzuführen.
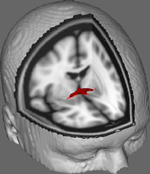
Abbildung 1. Gemittelte Tracking-Ergebnisse von 10 Kontrollteilnehmern, die einem T1-gewichteten Standard-MRT-Bild mit 2 mm isotropen Voxeln als anatomische Referenz überlagert wurden.
Gemittelte Tracking-Ergebnisse von allen 10 Kontrollteilnehmern wurden dann einem T1-gewichteten Standard-MRT-Bild mit 2 mm isotropen Voxeln als anatomische Referenz überlagert. Das mittlere FA-Verhältnis der Gruppe betrug 0,3146 (SD = 0,0472). Diese Traktographie-Ergebnisse zeigen, dass die Struktur des AC aufgeklärt werden kann und daher Potenzial für weitere Studien hat, in denen Leistungswerte bei Verhaltensaufgaben mit FA-Werten korreliert werden, wobei eine größere Stichprobe verwendet wird.
Zusammenfassend lässt sich sagen, dass es derzeit nur wenig Literatur gibt, und von der verfügbaren Literatur gibt es keine, die den AC speziell mit Prozessen im Zusammenhang mit dem AAA-Modell in Verbindung bringt, das im Mittelpunkt unseres früheren Zielartikels steht (Franz, 2012). Mit einem ähnlichen Verfahren wie dem hier vorgeschlagenen ist es möglich, größere Stichproben der Normalbevölkerung zu testen und die FA-Werte der DTI-Ergebnisse (zum Beispiel) mit einer spezifischen abhängigen Variable zu korrelieren, die aus einer eigens entwickelten Aufgabe der Handlungsaufmerksamkeit gewonnen wird (Verfahren, die in unserem Labor entwickelt werden).
Erklärung zu Interessenkonflikten
Die Autoren erklären, dass die Forschung in Abwesenheit jeglicher kommerzieller oder finanzieller Beziehungen durchgeführt wurde, die als potenzieller Interessenkonflikt ausgelegt werden könnten.
Danksagungen
Wir danken dem Human Brain Project und dem National Research Resource Center (USA) sowie der John Hopkins University für die Genehmigung zur öffentlichen Nutzung ihrer DTI-Daten. Wir sind auch dankbar für die großzügige Unterstützung durch den Marsden Fund of the Royal Society of New Zealand (an Elizabeth A. Franz).
Corballis, M. (1995). Visual integration in the split brain. Neuropsychologia 33, 937-959.
Pubmed Abstract | Pubmed Full Text
Franz, E. A. (2012). Die Zuweisung von Aufmerksamkeit zum Erlernen zielgerichteter Handlungen: ein kognitionsneurowissenschaftlicher Rahmen mit Fokus auf die Basalganglien. Front. Psychol. 3:535. doi: 10.3389/fpsyg.2012.00535
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Heath, C., and Jones, E. (1971). Interhemisphärische Bahnen beim Fehlen eines Corpus Callosum. An experimental study of commissural connexions in the marsupial phalanger. J. Anat. 109, 253-270.
Pubmed Abstract | Pubmed Full Text
Schreibe einen Kommentar