Superior olivary nucleus
On november 12, 2021 by admin| Hjerne: Superior olivary complex | ||||
|---|---|---|---|---|
 |
||||
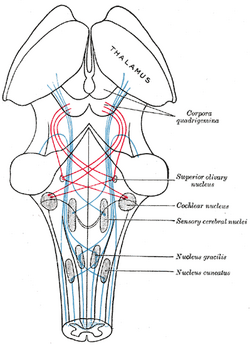
| Skema, der viser forløbet af lemniscusfibrene; medial lemniscus i blåt, lateral i rødt. (Superior olivary nucleus er mærket i midten til højre.) | ||||
| ] | ||||
| Latin | Latin | nucleus olivaris superior | ||
| Grays | subjekt #187 787 787 | |||
| Del af | ||||
| Komponenter | ||||
| Arterie | ||||
| Vein | ||||
| BrainInfo/UW | {{{{BrainInfoType}}}-565 | |||
| MeSH | ||||
Det overlegne olivariekompleks (eller SOC eller superior oliven) er en lille masse af grå substans beliggende på den dorsale overflade af den laterale del af trapezlegemet. Det er lille hos mennesker, men veludviklet hos visse dyr, og det udviser en lignende struktur som den nedre olivariekerne og er placeret umiddelbart over denne. Nogle af fibrene i trapezlegemet ender ved at arborisere omkring cellerne i denne kerne, mens andre udspringer fra disse celler.
Fysiologi
Den overlegne olivariekerne spiller en række roller i hørelsen. Den mediale superior olivenkerne (MSO) er en specialiseret kerne, som menes at måle tidsforskellen mellem lydenes ankomst mellem ørerne (den interaurale tidsforskel eller ITD). ITD er et vigtigt fingerpeg til bestemmelse af azimut for lavfrekvente lyde, dvs. til lokalisering af dem i det azimutale plan – deres grad til venstre eller højre.
Den laterale superior olive (LSO) menes at være involveret i måling af niveauforskellen af lyde mellem ørerne (den interaurale niveauforskel eller ILD). ILD er et andet vigtigt stikord til bestemmelse af azimut af højfrekvente lyde.
Forhold til det auditive system
Det overlegne olivariekompleks er generelt placeret i pons, men hos mennesker strækker det sig fra den rostrale medulla til den midterste pons og modtager projektioner overvejende fra den anteroventrale cochlear nucleus via den ventrale akustiske stria, selv om den posteroventrale nucleus også projicerer dertil via den mellemliggende akustiske stria. Det er på dette sted, at de første store binaurale interaktioner finder sted.
Primære kerner
Det overlegne olivariekompleks er opdelt i tre primære kerner, MSO, LSO og den mediale kerne af trapezlegemet, og flere mindre periolivære kerner. Disse tre kerner er de mest studerede og derfor bedst forstået. Typisk anses de for at danne den opadgående azimutale lokaliseringsvej.
Medial superior olive (MSO)
Den mediale superior olive menes at hjælpe med at lokalisere azimutten af en lyd, dvs. vinklen til venstre eller højre, hvor lydkilden er placeret. Ens første instinkt kan være at tro, at denne kerne omfatter vertikale oplysninger, men dette er ikke tilfældet; de oplysninger, der behandles i MSO, er begrænset til det horisontale plan. De fusiforme celler projicerer ikke til noget i niveau med pons og kommer først i spil ved den nedre colliculus inferior. Der er kun horisontale data til stede, men de kommer fra to forskellige ørekilder, hvilket hjælper med at lokalisere lyden på azimut-aksen. Den måde, hvorpå den overlegne oliven gør dette, er ved at måle forskellene i tid mellem to øresignaler, der registrerer den samme stimulus. Det tager ca. 700 μs at bevæge sig rundt om hovedet, og det antages, at den mediale superior olive er i stand til at registrere dette. Faktisk er det observeret, at mennesker kan registrere interaurale forskelle ned til 10 mikrosekunder. Kernen er tonotopisk organiseret, men den azimutale receptive feltprojektion er “højst sandsynligt et komplekst, ikke-lineært kort”.
Projektionerne fra den medial superior olive ender tæt i den ipsilaterale centrale kerne af inferior colliculus. Størstedelen af disse axoner anses for at være “rundformede” eller type R. Disse R-axoner er for det meste glutamatergiske og indeholder runde synaptiske vesikler og danner asymmetriske synaptiske junktioner.
- Dette er den største af kernerne og indeholder hos mennesker ca. 15.500 neuroner .
- Hver MSO modtager lavfrekvente bilaterale input fra højre og venstre AVCN.
- Udgangen er til den ipsilaterale laterale lemniscus og i sidste ende til den nedre colliculus.
- Den MSO reagerer bedre på binaurale stimuli.
- Denne hovedfunktion omfatter detektion af forskelle i ankomsttidspunktet for lyde til de to ører, hvilket er en del af lokaliseringsprocessen (ITD).
Lateral superior olive (LSO)
Denne olive har lignende funktioner som den mediale superior olive, men anvender intensitet til at lokalisere lydkilden. Det er den del af hjernestammen, der betegner den kraftigere lyd fra venstre øre som værende i venstre side. Den laterale olivenolvie modtager input fra begge cochlear nuclei, fra ipsilaterale AVCN sfæriske buskede celler og contralaterale AVCN globulære buskede celler via den ipsilaterale MNTB. MNTB-hovedcellerne er glycinergiske, hvilket giver en kilde til hæmning af LSO og danner grundlaget for ILD-følsomhed. Projektioner fra begge cochleære kerner er primært højfrekvente, og disse frekvenser er efterfølgende repræsenteret af størstedelen af LSO-neuronerne (>2/3 over 2-3 kHz hos kat). Yderligere input stammer fra den ipsilaterale LNTB (glycinergisk, se nedenfor), som giver hæmmende information fra den ipsilaterale cochlear nucleus. Et andet muligvis hæmmende input stammer fra ipsilaterale AVCN ikke-sfæriske celler. Disse celler er enten kugleformede buskede eller multipolære (stellate). Et af disse to input kan danne grundlag for ipsilateral hæmning, der ses i responskort, som flankerer den primære excitation og skærper enhedens frekvensafstemning.
LSO projicerer bilateralt til den centrale kerne af den nedre colliculus inferior (ICC). Ipsilaterale projektioner er primært hæmmende (glycinergiske), og kontralaterale excitatoriske. Yderligere projektionsmål omfatter de dorsale og ventrale kerner af den laterale lemniscus (DNLL & VNLL). De GABAergiske projektioner fra DNLL udgør den primære kilde til GABA i den auditive hjernestamme og projicerer bilateralt til ICC og til den kontralaterale DNLL. Disse konvergerende excitatoriske og inhiberende forbindelser kan virke til at mindske niveauafhængigheden af ILD-følsomheden i ICC sammenlignet med LSO.
Supplerende projektioner danner den laterale Olivocochlear Bundle (LOC), som innerverer cochleare indre hårceller. Disse projektioner menes at have en lang tidskonstant og fungerer til at normalisere det lydniveau, der registreres af hvert øre, for at hjælpe med lydlokalisering. Der er betydelige artsforskelle: LOC-projektionsneuroner er fordelt inden for LSO hos gnavere og omgiver LSO hos rovdyr (f.eks. kat).
Medial Nucleus of Trapezoid Body (MNTB)
- MNTB er den mindste del af SOC; tilstedeværelsen af MNTB i den menneskelige hjernestamme er tvivlsom.
- Hvert neuron modtager kun højfrekvente input via en Calyx af holdt fra den kontralaterale AVCN, og dets output projicerer til den ipsilaterale LSO.
- Der findes to responstyper: en ‘chopper-type’ svarende til spindelceller i AVCN og en primær type, som svarer til Bushy Cells i AVCN.
Periolivary Nuclei
SOC består af mellem seks og ni periolivary nuclei, afhængigt af den citerede forsker, der typisk navngives på baggrund af deres placering i forhold til de primære nuclei. Disse kerner omgiver hver af de primære kerner og bidrager til både det opadgående og det nedadgående auditive system. Disse kerner udgør også kilden til det olivocochleære bundt, som innerverer cochlea. Hos marsvinet er de opstigende projektioner til de nedre colliculi primært ipsilaterale (>80%), og den største enkeltkilde kommer fra SPON. Også de ventrale kerner (RPO, VMPO, AVPO, & VNTB) er næsten udelukkende ipsilaterale, mens de resterende kerner projicerer bilateralt.
| Navn | Kat | Guinea Pig | Rat | Mus | |||
|---|---|---|---|---|---|---|---|
| LSO | X | X | X | X | X | ||
| MSO | X | X | X | X | |||
| MNTB | X | X | X | X | X | X | X |
| LNTB | X | X | “LVPO” | X | |||
| ALPO | X | X | X | ||||
| PVPO | X | X | |||||
| PPO | X | X | “CPO” | ||||
| VLPO | X | ||||||
| DPO | X | X | X | ||||
| DLPO | X | X | X | ||||
| VTB | X | X | “MVPO” | X | |||
| AVPO | X | ||||||
| VMPO | X | X | |||||
| RPO | X | X | |||||
| SPN | “DMPO” | X | X | X | X |
Ventral Nucleus of Trapezoid Body (VNTB)
- VNTB er en lille kerne, der er placeret lateralt i forhold til MNTB, og ventral til MSO.
- Med en heterogen population af celler projicerer denne kerne til mange auditive kerner og danner det mediale olivocochleare bundle (MOC), som innerverer cochleare ydre hårceller . Disse celler indeholder elektromotilfibre og fungerer som mekaniske forstærkere/afdæmpere i cochlea.
- Kernen projicerer til begge IC, uden at der er nogen celler, der projicerer bilateralt.
Lateral Nucleus of the Trapezoid Body (LNTB)
- Lokaliseret ventral til LSO
- AVCN sfæriske buskede celler projicerer kollateraler bilateralt, og globulære buskede celler projicerer kollateraler ipsilateralt til LNTB-neuroner.
- Cellerne er immunoreaktive for glycin og er retrograd mærket efter injektion af tritieret glycin i LSO
- Kernen projicerer til både IC, med få celler, der projicerer bilateralt, samt til den ipsilaterale LSO.
- Store multipolære celler projicerer til den cochleare kerne, men ikke til IC, hos både kat og marsvin.
- Input sker ofte via endbulber af holdt, hvilket giver en meget hurtig signaltransduktion.
Superior Periolivary Nucleus (SPON) (Dorsomedial Periolivary Nucleus (DMPO))
- Lokaliseret direkte dorsalt i forhold til MNTB
- I rotte er SPON en homogen GABAerg kerne. Disse tonotopisk organiserede neuroner modtager excitatoriske input fra octopus og multipolære celler i den contralaterale ventrale cochlear nucleus, et glycinergisk (hæmmende) input fra den ipsilaterale MNTB, et ukendt GABAergisk (hæmmende) input, og projicerer til den ipsilaterale ICC. De fleste neuroner reagerer kun ved offset af en stimulus, kan faselåses til AM-stimuli på op til 200 Hz og kan danne grundlag for varighedsselektivitet i ICC. SPON-neuroner modtager ikke nedadgående input fra IC, og de projicerer ikke til cochlea eller cochlear nucleus, som mange periolivære kerner gør.
- I marsvin projicerer runde til ovale multipolære celler til begge IC, med mange celler, der projicerer bilateralt. De mere aflange celler, der projicerer til den cochleære kerne, projicerer ikke til ICC. Der synes at være to populationer af celler, en, der projicerer ipsilateralt, og en, der projicerer bilateralt.
- De fleste oplysninger var kommet fra gnaver-SPON på grund af kernenes fremtrædende størrelse hos disse arter, og der er kun foretaget meget få undersøgelser af DMPO hos katte, og ingen af dem var omfattende.
Dorsal Periolivary Nucleus (DPO)
- Lokaliseret dorsalt og medial til LSO
- Indeholder både EE-enheder (exciteret af begge ører) og E0-enheder (kun exciteret af det kontralaterale øre).
- Neuroner er tonotopisk organiserede og højfrekvente.
- Måtte tilhøre en enkelt kerne sammen med DLPO
- Kernen projicerer til begge IC, uden celler, der projicerer bilateralt.
Dorsolateral Periolivary Nucleus (DLPO)
- Lokaliseret dorsalt og lateralt i forhold til LSO
- Indeholder både EE (exciteret af begge ører) og E0 (kun exciteret af det kontralaterale øre) enheder.
- Neuroner er tonotopisk organiserede og lavfrekvente.
- Måtte tilhøre en enkelt kerne sammen med DPO
- Kernen projicerer til begge IC, med få celler, der projicerer bilateralt.
Ventrolateral Periolivary Nucleus (VLPO)
- Lokaliseret ventral til og inden for den ventrale hillus af LSO
- Indeholder både EI (exciteret af contralaterale og inhiberet af ipsilaterale øre) og E0 (kun exciteret af det contralaterale øre) enheder.
- Neuroner er tonotopisk organiserede og højfrekvente.
Anterolateral Periolivary Nucleus (ALPO)
- Kernen projicerer til begge IC, uden at der er nogen celler, der projicerer bilateralt.
- Store multipolære celler projicerer til den cochleære kerne, men ikke til IC, hos både kat og marsvin.
Ventromedial Periolivary Nucleus (VMPO)
- Lokaliseret mellem MSO og MNTB.
- Sender projektioner til ICC bilateralt.
- Kernen projicerer til begge IC, uden at der er nogen celler, der projicerer bilateralt.
Rostral Periolivary Nucleus (RPO) (Anterior Periolivary Nucleus (APO))
- Lokaliseret mellem den rostrale pol af MSO og VNLL
Caudal Periolivary Nucleus (CPO) (Posterior Periolivary Nucleus (PPO))
- Lokaliseret mellem den caudale pol af MSO og ansigtskernen (7N)
Posteroventral Periolivary Nucleus (PVPO)
- Kernen projicerer til begge IC, uden at der er nogen celler, der projicerer bilateralt.
Patofysiologi
Ved en obduktion af en 21-årig kvinde med autisme, epilepsi og mental retardering blev der fundet en næsten fuldstændig fravær af den overlegne oliven.
Se også
- Olivariallegeme
- Kulesza RJ, Cytoarchitecture of the human superior olivary complex: Medial and lateral superior olive. Hearing Research 225(2007) 80-90
- 2,0 2,1 Oliver DL, et al. Axonal projections from the lateral and medial superior olive to the inferior colliculus of the cat: a study using electron microscopic autoradiography. J Comp Neurol. 1995 Sep 11;360(1):17-32 Citerer fejl: Invalid
<ref>tag; name “R4” defined multiple times with different content - Cajal, S. R. Y. and L. Azoulay (1909). Histologie du système nerveux de l’homme et des vertébrés. Paris, Maloine.
- 4.0 4.1 Kandel, et al Principles of Neuroscience. Fjerde udgave. s. 591-624. Copyright 2000, af McGraw-Hill Co. Citerer fejl: Ugyldigt
<ref>tag; navnet “R2” defineret flere gange med forskelligt indhold - Kulesza, R. J., Jr. (2007). “Cytoarkitektur af det menneskelige overlegne olivariekompleks: medial og lateral overlegne oliven.” Hear Res 225(1-2): 80-90.
- Tsuchitani, C. og J. C. Boudreau (1967). “Kodning af stimulusfrekvens og intensitet af S-segmentceller i kattens superior olive.” J Acoust Soc Am 42(4): 794-805.
- 8.0 8.1 8.1 8.2 Citerer fejl: Invalid
<ref>tag; name “R10” defineret flere gange med forskelligt indhold - Wu, S. H. og J. B. Kelly (1994). “Fysiologisk bevis for ipsilateral hæmning i lateral superior oliven: synaptiske reaktioner i musens hjerneskive.” Hear Res 73(1): 57-64.
- Brownell, W. E., P. B. Manis, et al. (1979). “Ipsilaterale inhiberende responser i kattens lateral superior oliven.” Brain Res 177(1): 189-93.
- Darrow, K. N., S. F. Maison, et al. (2006). “Cochlear efferent feedback balancerer interaural følsomhed.” Nat Neurosci 9(12): 1474-6.
- Warr, W. B. og J. J. Guinan, Jr. (1979). “Efferent innervation af cortiorganet: to separate systemer.” Brain Res 173(1): 152-5.
- 13.0 13.1 Schofield, B. R. og N. B. Cant (1991). “Organisering af det overlegne olivariekompleks hos marsvinet. I. Cytoarkitektur, cytokromoxidase histokemi og dendritisk morfologi.” J Comp Neurol 314(4): 645-70. Citerer fejl: Invalid
<ref>tag; name “R7” defined multiple times with different content - 14.0 14.1 14.2 14.3 14.4 14.4 14.5 14.6 14.7 14.8 Illing, R. B., K. S. Kraus, et al. (2000). “Plasticitet i det overlegne olivariekompleks.” Microsc Res Tech 51(4): 364-81. Citerer fejl: Cite error: Cite error: Ugyldigt
<ref>tag; navnet “R5” defineret flere gange med forskelligt indhold Cite error: Invalid<ref>tag; name “R5” defined multiple times with different content Ugyldigt<ref>tag; navnet “R5” defineret flere gange med forskelligt indhold Cite error: Ugyldigt<ref>tag; navnet “R5” defineret flere gange med forskelligt indhold Cite error: Ugyldigt<ref>tag; navnet “R5” defineret flere gange med forskelligt indhold Cite error: Ugyldigt<ref>tag; navnet “R5” defineret flere gange med forskelligt indhold Cite error: Ugyldigt<ref>tag; navnet “R5” defineret flere gange med forskelligt indhold Cite error: Ugyldigt<ref>tag; navnet “R5” defineret flere gange med forskelligt indhold Cite error: Ugyldigt<ref>tag; navnet “R5” defineret flere gange med forskelligt indhold Cite error: Ugyldigt<ref>tag; navnet “R5” defineret flere gange med forskelligt indhold Cite error: Invalid<ref>tag; name “R5” defined multiple times with different content Cite error: Invalid<ref>tag; name “R5” defined multiple times with different content Cite error: Invalid<ref>tag; name “R5” defined multiple times with different content Cite error: Invalid<ref>tag; name “R5” defined multiple times with different content - Warr, W. B. and J. E. Beck (1996). “Flere projektioner fra den ventrale kerne af trapezlegemet hos rotte.” Hear Res 93(1-2): 83-101.
- 16.00 16.01 16.01 16.02 16.03 16.04 16.04 16.05 16.06 16.06 16.07 16.08 16.09 16.09 16.10 Schofield, B. R. og N. B. Cant (1992). “Organisering af det overlegne olivariekompleks hos marsvinet: II. Projektionsmønstre fra de periolivære kerner til den nedre colliculus.” J Comp Neurol 317(4): 438-55. Cite error: Cite error: Cite error: Ugyldigt
<ref>tag; navnet “R6” defineret flere gange med forskelligt indhold Cite error: Cite error: Ugyldigt<ref>tag; navn “R6” defineret flere gange med forskelligt indhold Cite error: Ugyldigt<ref>tag; navn “R6” defineret flere gange med forskelligt indhold Ugyldigt<ref>tag; navn “R6” defineret flere gange med forskelligt indhold Cite error: Ugyldigt<ref>tag; navn “R6” defineret flere gange med forskelligt indhold Ugyldigt<ref>tag; navnet “R6” defineret flere gange med forskelligt indhold Cite error: Ugyldigt<ref>tag; navnet “R6” defineret flere gange med forskelligt indhold Cite error: Ugyldigt<ref>tag; navn “R6” defineret flere gange med forskelligt indhold Cite error: Ugyldigt<ref>tag; navn “R6” defineret flere gange med forskelligt indhold Cite error: Ugyldigt<ref>tag; navn “R6” defineret flere gange med forskelligt indhold Cite error: Ugyldigt<ref>tag; navn “R6” defineret flere gange med forskelligt indhold Cite error: Ugyldigt<ref>tag; navn “R6” defineret flere gange med forskelligt indhold Cite error: Ugyldigt<ref>tag; navn “R6” defineret flere gange med forskelligt indhold Invalid<ref>tag; name “R6” defined multiple times with different content Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content - Smith, P. H., P. X. Joris, et al. (1993). “Projektioner af fysiologisk karakteriserede sfæriske buskcelleaxoner fra kokkekernen i kattens cochlear nucleus: bevis for forsinkelseslinjer til den mediale superior oliven.” J Comp Neurol 331(2): 245-60.
- Wenthold, R. J., D. Huie, et al. (1987). “Glycin immunoreaktivitet lokaliseret i den cochleare kerne og det overlegne olivariekompleks.” Neuroscience 22(3): 897-912.
- 19.0 19.1 Adams, J. C. (1983). “Cytologi af periolivære celler og organiseringen af deres projektioner hos katten.” J Comp Neurol 215(3): 275-89. Cite error: Invalid
<ref>tag; name “R8” defined multiple times with different content - Friauf, E. and J. Ostwald (1988). “Divergerende projektioner af fysiologisk karakteriserede neuroner i rotte ventrale cochlear nucleus-neuroner som vist ved intra-akonal injektion af peberrodsperoxidase.” Exp Brain Res 73(2): 263-84.
- Kulesza, R. J., Jr. og A. S. Berrebi (2000). “Superior paraolivær kerne hos rotten er en GABAerg kerne.” J Assoc Res Otolaryngol 1(4): 255-69.
- Kulesza, R. J., Jr., G. A. Spirou, et al. (2003). “Fysiologiske responsegenskaber af neuroner i den overlegne paraolivære kerne hos rotte.” J Neurophysiol 89(4): 2299-312.
- WHITE JS, WARR WB. Den dobbelte oprindelse af det olivocochleære bundt i albino-rotte. J. Comp. Neurol. 219:203-214, 1983.
- Guinan, J. J. J., Jr., S. S. Guinan, et al. (1972). “Enkelte auditive enheder i det overlegne olivariekompleks. I. Reaktioner på lyde og klassifikationer baseret på fysiologiske egenskaber.” Int J Neurosci 4: 101-20.
- Tsuchitani, C. (1977). “Funktionel organisation af laterale cellegrupper i kattens overlegne olivariekompleks.” J Neurophysiol 40(2): 296-318.
- Rodier PM, Ingram JL, Tisdale B, Nelson S, Romano J (1996). Embryologisk oprindelse for autisme: udviklingsmæssige anomalier i kranienervens motoriske kerner. J Comp Neurol 370 (2): 247-61.
- MedEd at Loyola Neuro/frames/nlBSs/nl27fr.htm
- Image at med.utah.edu (pick 9b. Pons)
Denne artikel var oprindeligt baseret på en post fra en public domain-udgave af Gray’s Anatomy. Som sådan kan nogle af de heri indeholdte oplysninger være forældede. Rediger venligst artiklen, hvis dette er tilfældet, og du er velkommen til at fjerne denne meddelelse, når den ikke længere er relevant.
|
v-d-e
Hjernen: rhombencephalon (baghjerne) |
||||||
|---|---|---|---|---|---|---|
| Metencephalon/ pons |
|
Dorsal/
(tegmentum) |
Ventral/
(base) |
Raphe/retikulær
|
||
| Myelencephalon/ medulla |
|
Dorsal
|
Ventral
|
Raphe/reticular
|
||
|
v-d-e
Sansesystem: Auditive og vestibulære systemer (TA A15.3, GA 10.1029) |
||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Udre øre |
Pinna (Helix, Antihelix, Tragus, Antitragus, Incisura anterior auris, Earlobe) – Øregang – Aurikulære muskler Earicrum (Umbo, Pars flaccida) |
|||||||||||||
| Mellemøre |
|
|||||||||||||
| Det indre øre/ (membranøs labyrint, benet labyrint) |
|
|||||||||||||
| {| class=”navbox collapsible nowraplinks” style=”margin:auto;” | ||||||||||||||
|
—
|
||||||||||||||
|
||||||||||||||
|}
Denne side bruger indhold med Creative Commons-licens fra Wikipedia (se forfattere).
Skriv et svar