Shark Week: Hvad spiste Megalodon?
On december 13, 2021 by adminGuest Author: Jack Cooper, MSc
UoB Graduate / PhD Student, University of Swansea
Som vi har set indtil nu, har der været ændringer og opdateringer af den videnskabelige konsensus om Megalodons taksonomiske tilknytning og maksimale kropsstørrelse. Dens yndlingssnack er dog noget, som der er universel enighed om i det videnskabelige samfund. Som en anden sjældenhed er det noget, der normalt er skildret med ret god nøjagtighed i fiktion. Tillad mig at præsentere en haj, der spiste hvaler – rigtige hvaler!
Mange af nutidens store makropredatorhajer er kendt for at spise havpattedyr, især vores gamle ven, den store hvide haj. Nogle af de mest ikoniske fotografier af denne haj omfatter den, der kaster sig op af vandet med sit seneste offer mellem kæberne (fig. 1). Unge hvide hajer spiser dog ikke sæler lige fra starten. I stedet starter de med at spise fisk og mindre hajer . Hvis vi igen ser på Gatun-formationen i Panama, finder vi masser af fossile hajer, rokker og otolitter (strukturer fra fiskenes indre ører) samt et væld af hvirvelløse dyr som bløddyr .
Det, vi ikke finder, er imidlertid fossile havpattedyr. Dette synes at indikere, at Megalodon-ungerne også spiste fisk og mindre hajer i stedet for umiddelbart at gå efter havpattedyr. Dette er noget, der kaldes et ontogenetisk skift i kosten, hvilket betyder, at hajen ændrer sin foretrukne fødekilde, efterhånden som den vokser. Ved at spise større føde, og sandsynligvis meget af den, da den nåede sin voksenstørrelse, ville Megalodon have været i stand til at opretholde de metaboliske krav, der var forbundet med at være så stor. Det kunne den have gjort ved at spise ikke kun hvaler, men stort set alt, hvad den kunne støde på.

I betragtning af hvor meget mad Megalodon skulle have spist for at retfærdiggøre sin enorme størrelse, var den næsten helt sikkert en opportunistisk æder. De fossile optegnelser er vores bedste guide her. Det skyldes tilstedeværelsen af sporfossiler, som er geologiske optegnelser af biologisk aktivitet. Det kan være alt fra fodspor fra dinosaurer til ridser på knogler, der viser, at noget har bidt det dyr, som knoglen tilhørte. Og Megalodon efterlod et par af disse sporfossiler, som giver vigtige beviser på dens drab.
Der er fundet flere fossiler tilhørende en række hvaler med bemærkelsesværdige bidemærker på. Et velciteret eksempel stammer fra Yorktown-formationen i Maryland, hvor flere hvalknogler havde bidemærker, der tydeligvis var lavet af store savtakkede tænder . Faktisk er der nogle gange fundet Megalodon-tænder direkte forbundet med disse knogler.
Hajer mister og udskifter tænder tusindvis af gange i løbet af deres liv, og derfor mister hajer ofte tænder, når de foretager et voldsomt drab. Dette er en væsentlig årsag til, at hajtænder er så almindelige fossiler. Tilstedeværelsen af hajtænder lige ved siden af et ridset hvalben tyder på, at der var fodring i gang. Men noget, som palæontologer altid bør overveje, når der findes sådanne fossiler, er: Var det faktisk et rovdyr? Eller var det måske en ådselshugst?
Sådselshugst er en almindelig fødeadfærd hos moderne makropredatoriske hajer som f.eks. storhvaler (fig. 2) og tigerhajer . Når disse hajer støder på døde hvaler, der flyder på overfladen, er det en buffet, hvor man kan spise alt, hvad man vil. Det er kritisk, at ådselsfiskeri ikke kræver nogen energiudgift, hvilket kan gå til spilde ved et mislykket rovdyrforsøg. I betragtning af at Megalodon var nødt til at kunne opretholde sin enorme størrelse, ville det ikke give meget mening at give afkald på et gratis måltid, hvis den stødte på en død hval. Derfor vil de fleste af os, hvis vi tager udgangspunkt i, hvordan store moderne hajer spiser, være villige til at satse på, at Megalodon formentlig spiste, når han fik chancen.

Dette sagt, er der et særligt bemærkelsesværdigt fossil, der implicerer Megalodon i et rovdyrforsøg. Og det er afgørende, at vi ved det primært, fordi forsøget mislykkedes. En artikel, der blev offentliggjort i 2010, beskrev et delvist ribbenpliocæn-fossil tilhørende en hval fra Yorktown-formationen i North Carolina (jep, to Yorktown-formationer). Dette fossil viste også skader, der lignede bidemærker, men især syntes der at være et vævet knogledække over disse mærker. Forfatterne tolkede dette som et tegn på en infektion, der var opstået, mens dyret var ved at komme sig efter knogletraumer forårsaget af disse bidemærker. Disse mærker passede godt sammen med et savtakket bid, hvilket tyder på, at angriberen havde savtakkede tænder – præcis som Megalodon. Undersøgelsen tyder derfor på, at en Megalodon eller en anden stor haj havde angrebet det dyr, som fossilet stammede fra, men at offeret havde overlevet forsøget, blot for at dø af en infektion fra sin skade seks uger senere.
Selv for nylig blev de første fossile bidemærker, der nogensinde er fundet på den sydlige halvkugle og tilskrives Megalodon, fundet i Perus Pisco-formation . Måske var det meningen, da det er det samme sted, hvor Gordon Hubbell fandt sit enestående Carcharodon-fossil (omtalt i Q1). Det bidte materiale, der blev fundet her, omfattede kranie- og ribbenrester, hvilket tyder på, at hajen havde bidt sit mål fra siden. Det mest betydningsfulde er, at resterne for første gang blev identificeret til en enkelt art – denne art er Piscobalaena nana, en lille bardehval. Ligesom i de andre sporfossiler passede bidemærkerne godt sammen med store savtakkede tænder – et godt argument for at beskylde Megalodon for angrebet (fig. 3). Det viste sig derfor, at Megalodon også var i stand til at angribe mindre hvaler (hvad enten det var et rovdyr eller et ådsel), hvilket er meget i tråd med forslagene om, at den sandsynligvis også angreb delfiner, havskildpadder og andre mindre byttedyr .

Selv om rovdriftsstrategier er svære at fortolke ud fra fossiler, kan moderne hajer give os en idé om, hvordan Megalodon jagede. Dens bidekraft ville have været usædvanlig kraftig til at beskadige hvalknogler. En computermodel af et kranium af en hvidhaj har foreslået, at den har en bidkraft på over ca. 1,8 tons, hvilket giver den en af de stærkeste bidkræfter af alle levende dyr . I forlængelse heraf tyder denne undersøgelse på, at Megalodons bidekraft var endnu større, idet den anslås til ekstraordinært 108.514-182.201 N.
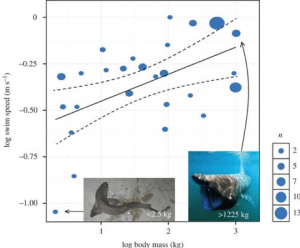
Videnskabsfolk har også gjort forsøg på at beregne Megalodons svømmehastighed, hvilket kan hjælpe os med at få en indikation af, hvordan den jagede. Hajer kan have en burst-hastighed, som er et hurtigt ryk med hurtig svømning, der typisk bruges til at gribe byttet. Men deres sædvanlige hastighed for deres afslappede svømmetag er det, vi kalder vedvarende svømmehastighed. Dette varierer fra art til art, men en undersøgelse under ledelse af David Jacoby kombinerede data om 26 arter fra 64 tidligere undersøgelser for at udvikle en model, der kunne forudsige et skaleringsforhold mellem vedvarende svømmehastighed og hajens stofskifte og kropsmasse. Deres model, der inkorporerede stofskiftet, afslørede en skaleringseksponent på 0,173, opsummeret som følgende ligning:
Svømmehastighed (ms ¯¹) ∝ Masse (kg)^0,173
Når de korrigerede dataene for fylogeni, fandt de imidlertid, at minimumssvømmehastighederne skalerede med kropsmassen ved den lidt lavere eksponent på 0,15. Ovenstående ligning forudsætter desuden en konstant på 1 for at passe til proportionerne, men dataene viste, at denne konstant faktisk var 0,266. De konstruerede derfor effektligningen:
Svømmehastighed (ms ¯¹) = 0,266 Masse (kg) ^0,15
Derudfra kan de beregne den vedvarende svømmehastighed for Megalodon afhængigt af dens masse. For eksempel ville den ~48.000 kg tunge Megalodon beregnet som massen af en 15,9 m lang haj i Gottfried’s arbejde afsløre en haj, der svømmer kontinuerligt med 1,34 m/s – ca. 4,8 km/t . Hvis vi anvender dette på en megalodon på 52.000 kg, får vi en kontinuerlig svømmehastighed på 1,36 m/s eller 4,9 km/t, mens hvis vi gik helt ekstremt til værks og simulerede en haj på 100.000 kg, ville denne model resultere i en svømmehastighed på 1,50 m/s eller 5,4 km/t. Denne model har vist sig at være meget effektiv, ikke kun til beregning af Megalodons svømmehastighed, men også til at levere en model for flere hajarter (fig. 4) .
Det er imidlertid den udbrudshastighed, som hajerne bruger til hurtigt at snuppe deres bytte. Hvidhajer udnytter dette til deres “breaching-adfærd”, hvor de driver sig selv op af vandet ved at ruske byttet op nedefra for at fange det ved overraskelse (fig. 1) . De kan gøre dette, fordi de er godt camouflerede på grund af deres modskygge. Så fordi Megalodon sandsynligvis også var modskygget og havde samme økologi og kropsstruktur som hvide hajer , rejser dette spørgsmålet: Var Megalodon også i stand til at bryde ud?
Måske. Hvidhajens udbrudshastighed er drevet af en fysiologisk tilpasning kaldet mesotermi . Dette er noget, som Megalodon for nylig er blevet foreslået også at have haft (vil blive diskuteret i detaljer i Q5). Mesoterme fisk har generelt en højere svømmehastighed ved vedvarende svømning og ved udbrud end ektoterme fisk , og dette er blevet brugt til at antyde, at Megalodon kan have haft en udbrudshastighed på op til 37,15 km/t .
Denne utroligt hurtige hastighed for en så stor haj kan meget vel være nok til at kunne bryde igennem. Ud over dette er den baskende haj (Cetorhinus maximus), som i dag er den næststørste haj, også blevet observeret bryde . Måske var den gigantiske megalodon virkelig i stand til at springe op af vandet, da den greb sit seneste måltid. Vi ved det bare ikke endnu, men min personlige og (lidt) professionelle mening er, at det meget vel kan have været muligt.
Med en så bred menu at vælge imellem var Megalodon et topprovdyr, som ikke havde nogen kendte rovdyr selv. Det betyder to vigtige ting. For det første havde den sandsynligvis hård konkurrence om sine byttedyr, hvilket i sidste ende kan have bidraget til dens udryddelse. For det andet ville Megalodon som topprovdyr have været af største betydning for sit økosystem, da den ville have holdt sine byttebestande i skak og dermed bevaret et sundt økosystem.
Det er derfor, at hajerne i dag er så vigtige for havmiljøerne. Tabet af en rovhaj, der er en af de vigtigste rovhajer, medfører irreversible kaskadevirkninger, der ændrer det samfund, som den efterlader . Som sådan er vores marine økosystemer som de er, delvist fordi Megalodon ikke længere er en del af dem. Og de enorme hvaler i de moderne oceaner kan hvile sig i ro og mag som følge heraf.
- Estrada JA, Rice AN, Natanson LJ & Skomal GB 2006. Anvendelse af isotopanalyse af ryghvirvler til rekonstruktion af ontogenetisk fødeøkologi hos hvidhajer. Ecology 87, 829-834.
- Pimiento C, Ehret DJ, MacFadden BJ & Hubbell G 2010. Gamle opvækstområder for den uddøde kæmpehaj Megalodon fra Miocæn i Panama. PLoS One 5, e10552.
- Pimiento C, González-Barba G, Ehret DJ, Hendy AJ, MacFadden BJ & Jaramillo C 2013. Hajer og rokker (Chondrichthyes, Elasmobranchii) fra den sene Miocæne Gatun-formation i Panama. J. Paleontol. 87, 755-774.
- Purdy RW 1996. Paleoøkologi af fossile hvide hajer. In: Great White Sharks: the biology of Carcharodon carcharias (eds. Klimley AP & Ainley DG), 67-78, Academic Press, San Diego.
- Aguilera OA, García L & Cozzuol MA 2008. Kæmpetandede hvide hajer og trofisk interaktion med hvaler fra den pliocæne caribiske Paraguaná-formation fra Pliocæn i Caribien. Paläontol. Z. 82, 204-208.
- Fallows C, Gallagher A. & Hammerschlag N 2013. Hvidhajer (Carcharodon carcharias), der æder hvaler, og dens potentielle rolle i yderligere udformning af økologien hos et topprovdyr. PLoS One, 8, e60797.
- Dudley SFJ, Anderson-Reade MD, Thompson GS & McMullen PB 2000. Samtidige plyndringer af et hvalkadaver af store hvide hajer, Carcharodon carcharias, og tigerhajer, Galeocerdo cuvier. Fish. Bull. 98, 646-649.
- Kallal RJ, Godfrey SJ & Ortner DJ 2010. Knoglereaktioner på et ribben fra en Pliocæn hval indikerer kortvarig overlevelse af en prædationshændelse. Int. J. Osteoarchaeol. 22, 253-260.
- Collareta A, Lambert O, Landini W, Di Celma C, Malinverno E, Varas-Malca R, Urbina M & Bianucci G 2017. Var den kæmpe uddøde haj Carcharocles megalodon målrettet mod små byttedyr? Bidemærker på marine pattedyrrester fra det sene Miocæn i Peru. Palaeogeogr. Palaeoclimatol. Palaeoecol. 469, 84-91.
- Wroe S, Huber DR, Lowry M, McHenry C, Moreno K, Clausen P, Ferrara TL, Cunningham E, Dean MN & Summers AP 2008. Tredimensional computeranalyse af hvidhajens kæbemekanik: Hvor hårdt kan en stor hvid haj bide? J. Zool. 276, 336-342.
- Jacoby DM, Siriwat P, Freeman R & Carbone C 2015. Er skalering af svømmehastighed hos hajer drevet af metabolisme? Biol. Lett. 11, 20150781.
- Gottfried MD, Compagno LJV & Bowman SC 1996. Størrelse og skeletanatomi hos den gigantiske “megatooth”-haj Carcharodon megalodon. In: Great White Sharks: the biology of Carcharodon carcharias (eds. Klimley AP & Ainley DG), 55-66, Academic Press, San Diego.
- Jacoby DM, Siriwat P, Freeman R & Carbone C 2016. Rettelse til “Er skaleringen af svømmehastighed hos hajer drevet af metabolisme? Biol. Lett. 12, 20160775.
- Watanabe YY, Payne NL, Semmens JM, Fox A & Huveneers C 2019. Svømmestrategier og energetik hos endoterme hvide hajer under fouragering. J. Exp. Biol. 222, jeb185603.
- Cooper JA, Pimiento C, Ferrón HG & Benton MJ (under tryk). Kropsdimensioner af den uddøde kæmpehaj Otodus megalodon: En 2D-rekonstruktion. Sci. Rep.
- Watanabe YY, Goldman KJ, Caselle JE, Chapman DD & Papastamatiou YP 2015. Sammenlignende analyser af dyresporingsdata afslører den økologiske betydning af endotermi hos fisk. Proc. Nat. Acad. Sci. 112, 6104-6109.
- Ferrón HG 2017. Regional endotermi som udløsende faktor for gigantisme hos nogle uddøde makropredatoriske hajer. PLoS One, 12, e0185185.
- Johnston EM, Halsey LG, Payne NL, Kock AA, Iosilevskii G, Whelan B & Houghton JD 2018. Latent kraft hos baskende hajer afsløret af ekstraordinære brydningsbegivenheder. Biol. Lett. 14, 20180537.
- Myers RA, Baum JK, Shepherd TD, Powers SP & Peterson CH 2007. Kaskadevirkninger af tabet af spidse rovhajer fra et kystnært hav. Science, 315, 1846-1850.
Udgivet af Rhys Charles
Skriv et svar