N-linked glycosylering
On december 18, 2021 by admin
Biosyntesen af N-linkede glykaner foregår via 3 hovedtrin:
- Syntese af dolichol-linked precursor oligosaccharid
- En bloc overførsel af precursor oligosaccharid til protein
- Bearbejdning af oligosaccharidet
Syntese, en bloc overførsel og indledende trimning af precursor oligosaccharidet sker i det endoplasmatiske reticulum (ER). Efterfølgende bearbejdning og modifikation af oligosakkaridkæden foregår i Golgi-apparatet.
Syntesen af glykoproteiner er således rumligt adskilt i forskellige cellekompartmenter. Derfor afhænger den type N-glykan, der syntetiseres, af dets tilgængelighed for de forskellige enzymer, der er til stede i disse cellekompartmenter.
Men på trods af forskelligheden syntetiseres alle N-glykaner gennem en fælles vej med en fælles kerneglykanstruktur.
Kerneglykanstrukturen består i det væsentlige af to N-acetylglucosamin- og tre mannoserester. Dette kerneglykan udarbejdes og modificeres derefter yderligere, hvilket resulterer i et varieret udvalg af N-glykanstrukturer.
Syntese af prækursoroligosaccharidRediger
Processen for N-bunden glykosylering starter med dannelsen af dolichol-bundet GlcNAc-sukker. Dolichol er et lipidmolekyle, der er sammensat af gentagne isopren-enheder. Dette molekyle findes fastgjort til ER’s membran. Sukkermolekyler er knyttet til dolichol gennem en pyrofosfatbinding (det ene fosfat var oprindeligt knyttet til dolichol, og det andet fosfat kom fra nukleotidsukkeret). Oligosakkaridkæden forlænges derefter gennem tilsætning af forskellige sukkermolekyler trinvis for at danne et forstadieoligosakkarid.
Samlingen af dette forstadieoligosakkarid sker i to faser: Fase I og II. Fase I finder sted på den cytoplasmatiske side af ER, og fase II finder sted på den luminale side af ER.
Det forstadiemolekyle, der er klar til at blive overført til et protein, består af 2 GlcNAc-, 9 mannose- og 3 glukosemolekyler.

|
|
| Fase II | |
er mandoserestdonor(dannelse : Dol-P + GDP-Man → Dol-P-Man + GDP) og Dol-P-Gluc er glukoserestdonor(dannelse : Dol-P + UDP-Glc → Dol-P-Glc + UDP).
|
|
Overførsel af glykan til proteinRediger
Når prækursoroligosakkaridet er dannet, overføres det færdige glykan derefter til det spirende polypeptid i lumenet af ER-membranen. Denne reaktion drives af den energi, der frigives fra spaltningen af pyrofosfatbindingen mellem dolichol-glykanmolekylet.Der er tre betingelser, der skal være opfyldt, før et glykan overføres til et spirende polypeptid:
- Asparagin skal være placeret i en specifik konsensussekvens i primærstrukturen (Asn-X-Ser eller Asn-X-Thr eller i sjældne tilfælde Asn-X-Cys).
- Asparagin skal være placeret hensigtsmæssigt i proteinets tredimensionelle struktur (sukkerstoffer er polære molekyler og skal derfor være knyttet til asparagin placeret på proteinets overflade og ikke begravet i proteinet)
- Asparagin skal findes i den luminale side af det endoplasmatiske retikulum, for at N-bunden glykosylering kan påbegyndes. Targetrester findes enten i sekretoriske proteiner eller i de regioner af transmembranproteiner, der vender ud mod lumenet.
Oligosaccharyltransferase er det enzym, der er ansvarlig for genkendelse af konsensussekvensen og overførsel af forløberglykanet til en polypeptidacceptor, som er ved at blive oversat i lumenet af det endoplasmatiske reticulum. N-linked glykosylering er derfor er er en co-translationel begivenhed
Behandling af glykanerRediger

N-glykanbehandling foregår i endoplasmatisk retikulum og Golgi-legemet. Den indledende trimning af prækursormolekylet sker i ER, og den efterfølgende behandling finder sted i Golgi.
Når det færdige glykan overføres til det spirende polypeptid, fjernes to glukoserester fra strukturen. Enzymer, der er kendt som glykosidaser, fjerner nogle sukkerrester. Disse enzymer kan bryde glykosidiske bindinger ved hjælp af et vandmolekyle. Disse enzymer er exoglykosidaser, da de kun arbejder på monosakkaridrester, der er placeret i den ikke-reducerende ende af glykanet. Dette indledende trimningstrin menes at fungere som et kvalitetskontroltrin i ER for at overvåge proteinfoldningen.
Når proteinet er foldet korrekt, fjernes to glukoserester af glucosidase I og II. Fjernelsen af den sidste tredje glukoserest signalerer, at glykoproteinet er klar til transit fra ER til cis-Golgi. ER mannosidase katalyserer fjernelsen af denne sidste glukose. Hvis proteinet imidlertid ikke er foldet korrekt, fjernes glukoseresterne ikke, og glykoproteinet kan derfor ikke forlade det endoplasmatiske reticulum. Et chaperonprotein (calnexin/calreticulin) binder sig til det udfoldede eller delvist foldede protein for at hjælpe med proteinfoldningen.
Det næste trin indebærer yderligere tilføjelse og fjernelse af sukkerrester i cis-Golgi. Disse modifikationer katalyseres af henholdsvis glykosyltransferaser og glykosidaser. I cis-Golgi fjerner en række mannosidaser nogle af eller alle de fire mannoserester i α-1,2-bindinger. Mens glykosyltransferaser i den mediale del af Golgi tilføjer sukkerrester til kerne-glykanstrukturen, hvilket giver anledning til de tre hovedtyper af glykaner: high mannose-, hybrid- og komplekse glykaner.
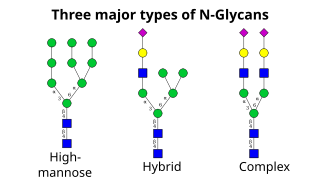
- Høj mannose er i bund og grund blot to N-acetylglucosaminer med mange mannoserester, ofte næsten lige så mange som der ses i forløber-oligosacchariderne, før det er knyttet til proteinet.
- Komplekse oligosakkarider hedder sådan, fordi de kan indeholde næsten et hvilket som helst antal af de andre typer saccharider, herunder flere end de oprindelige to N-acetylglucosaminer.
- Hybridoligosakkarider indeholder en mannoserest på den ene side af grenen, mens en N-acetylglucosamin på den anden side indleder en kompleks forgrening.
Rækkefølgen af tilføjelse af sukkerstoffer til de voksende glykan-kæder bestemmes af enzymernes substratspecificiteter og deres adgang til substratet, når de bevæger sig gennem sekretoriske veje. Således spiller organiseringen af dette maskineri i en celle en vigtig rolle for, hvilke glykaner der laves.
Enzymer i GolgiEdit
Golgi-enzymer spiller en vigtig rolle i bestemmelsen af syntesen af de forskellige typer af glykaner. Enzymernes virkningsrækkefølge afspejles i deres placering i Golgi-stakken:
| Enzymer | Lokalisering i Golgi |
|---|---|
| Mannosidase I | cis-Golgi |
| GlcNAc-transferaser | medial Golgi |
| Galactosyltransferase og Sialyltransferase | trans-Golgi |
I archaea og prokaryoterRediger
Lignende N-glycan biosynteseveje er fundet i prokaryoter og Archaea. Sammenlignet med eukaryoter synes den endelige glykanstruktur i eubakterier og archaea imidlertid ikke at afvige meget fra den oprindelige forløber, der er fremstillet i det endoplasmatiske reticulum. I eukaryoter modificeres det oprindelige oligosaccharidprækursor-oligosaccharid omfattende undervejs til celleoverfladen.
Skriv et svar