Hebbiansk teori
On december 14, 2021 by admin Vurdering | Biopsykologi | Komparativ | Kognitiv | Udvikling | Sprog | Individuelle forskelle | Personlighed | Filosofi | Social |
Metoder | Statistik | Klinisk | Pædagogisk | Industriel | Professionelle emner | Verdenspsykologi |
Kognitiv psykologi:Opmærksomhed – Beslutningstagning -Læring – Dømmekraft – Hukommelse – Motivation – Opfattelse – Ræsonnement -Tænkning -Tænkning -Kognitive processerKognition -OutlineIndex
Hebbian-teorien beskriver en grundlæggende mekanisme for synaptisk plasticitet, hvor en stigning i synaptisk effektivitet opstår som følge af den præsynaptiske celles gentagne og vedvarende stimulering af den postsynaptiske celle. Den blev introduceret af Donald Hebb i 1949 og kaldes også Hebbs regel, Hebbs postulat og celleforsamlingsteori og lyder:
Lad os antage, at vedholdenhed eller gentagelse af en reverberende aktivitet (eller “spor”) har en tendens til at fremkalde varige cellulære ændringer, der øger dens stabilitet…. Når en axon fra celle A er tæt nok på til at excitere en celle B og gentagne gange eller vedvarende deltager i affyringen af den, sker der en eller anden vækstproces eller metabolisk ændring i en eller begge celler, således at A’s effektivitet, som en af de celler, der affyrer B, øges.
Teorien opsummeres ofte som “celler, der fyrer sammen, kobler sig sammen”, selv om dette er en oversimplificering af nervesystemet, som ikke skal tages bogstaveligt, og som heller ikke nøjagtigt repræsenterer Hebbs oprindelige udsagn om ændringer i cellernes forbindelsesstyrke. Teorien fremkaldes ofte for at forklare visse former for associativ indlæring, hvor samtidig aktivering af celler fører til udpræget stigning i synaptisk styrke. En sådan læring er kendt som hebbisk læring.
Hebbiske engrammer og celleforsamlingsteori
Hebbisk teori vedrører, hvordan neuroner kan forbinde sig selv for at blive til engrammer. Hebbs teorier om celleforsamlingers form og funktion kan forstås ud fra følgende:
“Den generelle idé er gammel, at to celler eller systemer af celler, der gentagne gange er aktive på samme tid, vil have en tendens til at blive ‘associeret’, således at aktivitet i den ene letter aktivitet i den anden.” (Hebb 1949, s. 70) “Når en celle gentagne gange hjælper med at affyre en anden, udvikler den første celles axon synaptiske knopper (eller udvider dem, hvis de allerede findes) i kontakt med den anden celles soma.” (Hebb 1949, s. 63)
Gordon Allport fremsætter yderligere ideer vedrørende teorien om celleforsamling og dens rolle i dannelsen af engrammer, i tråd med begrebet auto-association, der beskrives således:
“Hvis input til et system forårsager, at det samme aktivitetsmønster opstår gentagne gange, vil det sæt af aktive elementer, der udgør dette mønster, blive stadig stærkere interassocieret. Det vil sige, at hvert element vil have en tendens til at tænde for hvert andet element og (med negative vægte) til at slukke for de elementer, der ikke indgår i mønsteret. Med andre ord vil mønstret som helhed blive “auto-associeret”. Vi kan kalde et indlært (auto-associeret) mønster for et engram.” (Hebb 1949, s. 44)
Hebbiansk teori har været det primære grundlag for den konventionelle opfattelse, at engrammer, når de analyseres fra et holistisk niveau, er neuronale net eller neurale netværk.
Arbejde i Eric Kandels laboratorium har givet beviser for inddragelse af hebbianske læringsmekanismer ved synapser i havsneglen Aplysia californica.
Eksperimenter på hebbiske synapsmodifikationsmekanismer ved synapserne i centralnervesystemet hos hvirveldyr er meget vanskeligere at kontrollere end eksperimenter med de relativt enkle synapser i det perifere nervesystem, der er undersøgt hos marine hvirvelløse dyr. En stor del af arbejdet med langvarige synaptiske ændringer mellem hvirveldyrs neuroner (som f.eks. langtidspotentiering) indebærer anvendelse af ikke-fysiologisk eksperimentel stimulering af hjerneceller. Nogle af de fysiologisk relevante mekanismer til ændring af synapser, der er blevet undersøgt i hvirveldyrhjerner, synes imidlertid at være eksempler på hebbiske processer. En sådan undersøgelse gennemgår resultater fra eksperimenter, der viser, at langvarige ændringer i synaptiske styrker kan induceres af fysiologisk relevant synaptisk aktivitet, der virker gennem både hebbiske og ikke-hebbiske mekanismer
Principper
Fra et synspunkt med kunstige neuroner og kunstige neurale netværk kan Hebbs princip beskrives som en metode til at bestemme, hvordan vægtene mellem modelneuroner skal ændres. Vægten mellem to neuroner øges, hvis de to neuroner aktiveres samtidig – og reduceres, hvis de aktiveres separat. Knuder, der har tendens til at være enten begge positive eller begge negative på samme tid, har stærke positive vægte, mens knuder, der har tendens til at være modsatrettede, har stærke negative vægte.
Dette oprindelige princip er måske den enkleste form for vægtvalg. Selv om det betyder, at det relativt let kan kodes ind i et computerprogram og bruges til at opdatere vægtene for et netværk, forbyder det også antallet af anvendelsesmuligheder for hebbisk læring. I dag henviser udtrykket hebbisk læring generelt til en form for matematisk abstraktion af det oprindelige princip, der blev foreslået af Hebb. I denne forstand indebærer hebbisk indlæring, at vægtene mellem indlæringsknuder justeres, således at hver vægt bedre repræsenterer forholdet mellem knuderne. Som sådan kan mange læringsmetoder betragtes som værende af en vis hebbisk karakter.
Det følgende er en formelmæssig beskrivelse af hebbisk læring: (Bemærk, at mange andre beskrivelser er mulige)
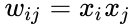
hvor 
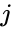
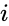
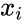
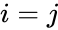
En anden formelmæssig beskrivelse er:
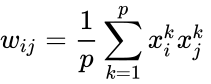
hvor 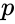

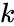
En variant af hebbisk læring, der tager hensyn til fænomener som blokering og mange andre neurale læringsfænomener, er Harry Klopfs matematiske model. Klopfs model gengiver en lang række biologiske fænomener og er desuden enkel at implementere.
Generalisering og stabilitet
Hebbs regel generaliseres ofte som
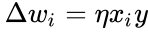
eller ændringen i den 
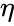
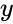
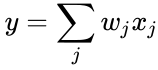
og i forrige afsnits forenkling antages både indlæringshastigheden og inputvægtene at være 1. Denne version af reglen er klart ustabil, da i ethvert netværk med et dominerende signal vil de synaptiske vægte stige eller falde eksponentielt. Det kan imidlertid påvises, at Hebbs regel er ustabil for enhver neuronmodel. Derfor anvender netværksmodeller af neuroner normalt andre indlæringsteorier som f.eks. BCM-teorien, Ojas regel eller den generaliserede hebbiske algoritme.
Se også
- Anti-Hebbian learning
- BCM theory
- Coincidence Detection in Neurobiology
- Dale’s principle
- Generalized Hebbian Algorithm
- Leabra
- Long-term potentiation
- Memory
- Metaplasticity
- Neurale netværk
- Oja learning rule
- Tetanic stimulation
- Spike-timing-afhængig plasticitet
- Synaptotropisk hypotese
- Shouval, Harel Hjernefysikken i hjernen. Det synaptiske grundlag for læring og hukommelse: En teoretisk tilgang. The University of Texas Health Science Center at Houston. URL tilgået den 2007-11-14.
Yderligere læsning
- Hebb, D.O. (1949), The organization of behavior, New York: Wiley
- Hebb, D.O. (1961). “Distinctive features of learning in the higher animal” J. F. Delafresnaye (Ed.) Brain Mechanisms and Learning, London: Oxford University Press.
- Hebb, D.O., og Penfield, W. (1940). Menneskets adfærd efter omfattende bilateral fjernelse fra frontallapperne. Archives of Neurology and Psychiatry 44: 421-436.
- Allport, D.A. (1985). “Distributed memory, modular systems and dysphasia” Newman, S.K. and Epstein, R. (Eds.) Current Perspectives in Dysphasia, Edinburgh: Churchill Livingstone.
- Bishop, C.M. (1995). Neural Networks for Pattern Recognition (Neurale netværk til mønstergenkendelse), Oxford: Oxford University Press.
- Paulsen, O., Sejnowski, T. J. (2000). Naturlige aktivitetsmønstre og langsigtet synaptisk plasticitet. Current opinion in neurobiology 10 (2): 172-179.
- Overblik
- Hebbian Learning tutorial (Del 1: Novelty Filtering, Del 2: PCA)
|
v-d-e
Neuroethologi |
||
|---|---|---|
| Begreber i neuroethologi |
Feedforward – Coincidence detector – Umwelt – Instinkt – Feature detektor – Central mønstergenerator (CPG) -NMDA-receptor – Lateral hæmning – Fast handlingsmønster – Kroghs princip-Hebbian teori- Lydlokalisering |
200px
|
| Historie om neuroethologi |
Theodore Holmes Bullock – Walter Heiligenberg -Niko Tinbergen- Konrad Lorenz- Eric Knudsen-Donald Griffin – Donald Kennedy – Karl von Frisch – Erich von Holst – Jörg-Peter Ewert |
|
| Metoder i neuroethologi |
Helcelle Patch Clamp |
|
| Modelsystemer i neuroethologi |
Dyrs ekkolokalisering – Vaggle Dance- Electric Fish – Vision in toads – Neuroethology of Frog Audition – Infrared sensing in snakes |
|
Denne side bruger indhold med Creative Commons-licens fra Wikipedia (se forfattere).
Skriv et svar