Bakteriecellers struktur og funktion
On oktober 10, 2021 by adminBakteriecellers struktur og funktion (side 5)
(Dette kapitel har 10 sider)
© Kenneth Todar, PhD
Cellevæg
Cellevæggen
Bakteriernes cellevægge fortjener særlig opmærksomhed af flere grunde:
1. De er en væsentlig struktur for levedygtighed, som beskrevet ovenfor.
2. De er sammensat af unikke komponenter, som ikke findes andre steder i naturen.
3. De er et af de vigtigste steder for angreb fra antibiotika.
4. De giver ligander til adhærens og receptorsteder for lægemidler eller virus.
5. De forårsager sygdomssymptomer hos dyr.
6. De giver mulighed for immunologisk skelnen og immunologisk variation mellem bakteriestammer.
De fleste procaryoter har en stiv cellevæg.Cellevæggen er en vigtig struktur, der beskytter celleprotoplasten mod mekanisk skade og mod osmotisk brud eller lysis.Prokaryoter lever normalt i relativt fortyndede miljøer, således at ophobningen af opløste stoffer i den prokaryote cellecytoplasma i høj grad overstiger den samlede koncentration i det ydre miljø. Det osmotiske tryk mod indersiden af plasmamembranen kan således svare til 10-25 atm. Da membranen er en skrøbelig, plastisk struktur, må den holdes tilbage af en ydre væg af porøst, stift materiale med høj trækstyrke. Et sådant materiale er murein, som er den allestedsnærværende bestanddel af bakterielle cellevægge.
Murein er en unik type petidoglykan, en polymer af disaccharider (glykan), der er tværbundet med korte kæder af aminosyrer (peptid). Der findes mange typer af peptidoglykan. Alle bakterielle peptidoglykaner indeholder N-acetylmuraminsyre, som er den definerende bestanddel af murein. Archaea’s cellevægge kan bestå af protein, polysaccharider eller peptidoglykanlignende molekyler, men de indeholder aldrig murein. Denne egenskab adskiller bakterierne fra arkæerne.
I de Gram-positive bakterier (de bakterier, der bevarer det lilla krystalviolette farvestof ved Gram-farvning) består cellevæggen af flere lag peptidoglykan. Løbende vinkelret på peptidoglykanlagene findes en gruppe af molekyler kaldet teichoiske syrer, som er unikke for den Gram-positive cellevæg (figur 14). 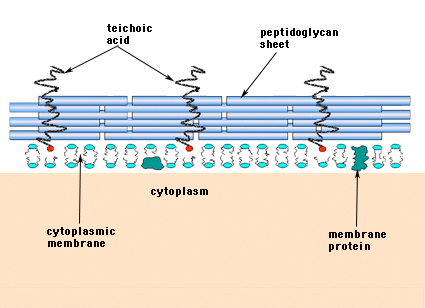
Figur 14. Strukturen af den Gram-positive bakteriers cellevæg. Væggen er forholdsvis tyk og består af mange lag peptidoglykan, der er gennemsat med teichoinsyrer, som løber vinkelret på peptidoglykanlagene.
I de gramnegative bakterier (som ikke tilbageholder krystalviolet) består cellevæggen af et enkelt lag af peptidoglykaner omgivet af en membranagtig struktur kaldet den ydre membran.Den ydre membran hos gramnegative bakterier indeholder altid en enkelt komponent, lipopolysaccharid (LPS eller endotoxin), som er giftig for dyr. Hos gramnegative bakterier betragtes den ydre membran normalt som en del af cellevæggen (figur 15).

Figur 15. Strukturen af den gramnegative cellevæg. Væggen er forholdsvis tynd og indeholder meget mindre peptidoglykan end den Gram-positive væg, og der er heller ingen teichoiske syrer. Den Gram-negative cellevæg består imidlertid af en ydre membran, som ligger uden for peptidoglykanlaget. Den ydre membran er fastgjort til peptidoglykanlaget af enunik gruppe af lipoproteinmolekyler.
I de Gram-positive bakterier er cellevæggen tyk (15-80anometer) og består af flere lag peptidoglykan. Hos de Gram-negative bakterier er cellevæggen forholdsvis tynd (10 nanometer) og består af et enkelt lag af peptidoglykaner omgivet af en ydre membran.
Peptidoglykanstrukturen og -arrangementet i E.coli er repræsentativt for alle Enterobacteriaceae samt for mange andre gramnegative bakterier. Glykanryggen består af alternative molekyler af N-acetylglucosamin (G) og N-acetylmuraminsyre (M), der er forbundet med en beta 1,4-glycosidbinding. 3-kulstoffet i N-acetylmuraminsyre (M) er substitueret med en lactylethergruppe, der stammer fra pyruvat. Lactylethergruppen forbinder glykanryggen med en peptidsidekæde, der indeholder L-alanin (L-ala), D-glutamat (D-glu), diaminopimelinsyre (DAP) og D-alanin (D-ala). MurNAc er unikt for bakterielle cellevægge, ligesom D-glu, DAP og D-ala. Muraminsyreunderenheden i E. coli er vist iFigur16 nedenfor.

Figur 16. Strukturen af muraminsyreunderenheden i peptidoglykanet fra Escherichia coli.Dette er den type murein, der findes i de fleste gramnegative bakterier. Glykanryggen er en gentagen polymer af to aminosukkerstoffer, N-acetylglucosamin (G) og N-acetylmuraminsyre (M). Til N-acetylmuraminsyre er knyttet etetrapeptid bestående af L-ala-D-glu-DAP-D-ala. b. Forkortet struktur af muraminsyreunderenheden. c. Nærliggende tetrapeptidsidekæder kan være forbundet med hinanden ved en interpeptidbinding mellem DAP på den ene kæde og D-ala på den anden. d. Molekylets polymere form.
Mureinstrenge er samlet i periplasmaet af ca. 10 muraminsyreunderenheder. Derefter forbindes strengene til et sammenhængende glykanmolekyle, der omslutter cellen.Hvor deres nærhed tillader det, kan de tetrapeptidkæder, der rager ud fra glykanryggen, tværbindes ved hjælp af en interpeptidbinding mellem en fri aminogruppe på DAP og en fri carboxygruppe på en nærliggende D-ala.Samlingen af peptidoglykan på ydersiden af plasmamembranen sker ved hjælp af en gruppe periplasmiske enzymer, som er transglycosylaser, transpeptidaser og carboxypeptidaser. Virkningsmekanismen for penicillin og beslægtede beta-lactam-antibiotika er at blokere transpeptidase- og carboxypeptidaseenzymerne under deres samling af murein-cellevæggen. Derfor siges beta-actam-antibiotika at “blokere cellevægssyntesen” i bakterierne.
Glykanryggen i peptidoglykanmolekylet kan spaltes af et enzym kaldet lysozym, der findes i animalsk serum, væv og sekreter og i det fagocytiske lysosom. Lysozymets funktion er at lyse bakterieceller som et konstitutivt forsvar mod bakterielle patogener. Nogle Gram-positive bakterier er meget følsomme over for lysysozyme, og enzymet er ret aktivt ved lave koncentrationer.Lakriske sekretioner (tårer) kan fortyndes i forholdet 1:40 000 og bevarer evnen til at kunne lyse visse bakterieceller. Gram-negative bakterier er mindre sårbare over for lysozymets angreb, fordi deres peptidoglykan er afskærmet af ydermembranen. Det nøjagtige sted for lysozymets spaltning er beta 1,4-bindingen mellem N-acetylmuraminsyre (M) og N-acetylglucosamin (G), således at den i figur 16(a) viste underenhed af duraminsyre er resultatet af lysozymets påvirkning af bakterielt peptidoglycan.
I Gram-positive bakterier findes der mangeforskellige peptidarrangementer blandt peptidoglycaner. Den bedst undersøgte er denurein af Staphylococcus aureus, der er vist i figur 17 nedenfor. I stedet for DAP (i E. coli) er diaminsyren L-lysin (L-lys), og i stedet for interpeptidbindingen (i Gram-negative) er der en interpeptidbro af aminosyrer, der forbinder en fri aminogruppe på lysin med en fri karboxygruppe på D-ala i en nærliggende tetrapeptidsidekæde. Dette arrangement giver tilsyneladende mulighed for hyppigere krydsbindinger mellem nærliggende tetrapeptidsidekæder. I S. aureus er interpeptidbroen en peptiddekoration bestående af 5 glycinmolekyler (kaldet en pentaglycinbro). Samlingen af interpeptidbroen i Gram-positivt murein hæmmes af beta-lactam-antibiotika på samme måde som interpeptidbindingen i Gram-negativt murein. Gram-positive bakterier er mere følsomme over for penicillin end Gram-negative bakterier, fordi peptidoglykanet ikke er beskyttet af en ydre membran, og fordi det er et hyppigere molekyle. I Gram-positive bakterier kan peptidoglykaner variere med hensyn til den aminosyre, der erstatter DAP eller L-L-lys i position 3 i tetrapeptidet, og med hensyn til den nøjagtige sammensætning af interpeptidbroen. Der findes mindst otte forskellige typer af peptidoglykaner i Gram-positive bakterier.
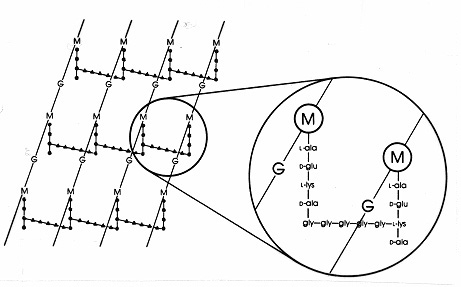
Figur 17. Skematisk diagram af peptidoglykanlaget hos Staphylococcus aureus. G = N-acetyl-glucosamin;M = N-acetyl-muraminsyre; L-ala = L-alanin; D-ala = D-alanin; D-glu= D-glutaminsyre; L-lys = L-lysin. Dette er en type murein, der findes i Gram-positive bakterier. I forhold til E. coli-peptidoglykanet (figur 7) er der L-lys i stedet for DAP (diaminopimelinsyre) i tetrapeptidet.L-lys’ frie aminogruppe er substitueret med et glycinpentapeptid (gly-gly-gly-gly-gly-gly-gly-gly-), som derefter bliver en interpeptidbro, der danner en forbindelse med en carboxygruppe fra D-ala i en tilstødende tetrapeptidsidekæde. Gram-positive peptidoglycaner adskiller sig fra art til art, hovedsagelig med hensyn til aminosyrerne i tredje position i tetrapeptidkæden og i aminosyresammensætningen af interpeptidbroen.
Gram-negative bakterier kan indeholde et enkeltlemonomolekylært lag murein i deres cellevægge, mens Gram-positive bakterier antages at have flere lag eller “indpakninger” af peptidoglycan. Tæt forbundet med lagene af peptidoglykan i Gram-positive bakterier er en gruppe af molekyler, der kaldes teichoiske syrer. Teikosyrer er lineære polymerer af polyglycerol eller polyribitol, der er substitueret med fosfater og nogle få aminosyrer og sukkerarter. Teichoesyrepolymererne er lejlighedsvis forankret i plasmamembranen (kaldet lipoteichoesyre, LTA) og tilsyneladende rettet udad vinkelret på peptidoglykanlagene. Man kender ikke funktionen af teichoesyre. De er afgørende for Gram-positive bakteriers levedygtighed i naturen. En idé er, at de udgør en kanal med regelmæssigt orienterede negative ladninger til at føre positivt ladede stoffer gennem det komplicerede peptidoglycan-netværk. En anden teori er, at teichoesyrer på en eller anden måde er involveret i regulering og samling af muraminsyre-underenheder på ydersiden af plasmamembranen. Der er eksempler, især i streptokokker, hvor teichoiske syrer er blevet inddraget i bakteriernes vedhæftning til vævsoverflader.
Kapitel fortsat
Forrige side
Skriv et svar