Týden se žraloky:
On 13 prosince, 2021 by adminHost Author: What Did Megalodon Eat? Jack Cooper, MSc
UoB Graduate / PhD Student, University of Swansea
Jak jsme dosud viděli, došlo ke změnám a aktualizacím vědeckého konsensu ohledně taxonomického zařazení megalodona a jeho maximální tělesné velikosti. Nicméně jeho oblíbená svačina je něco, na čem se vědecká komunita všeobecně shoduje. Další raritou je, že je to něco, co je obvykle s docela dobrou přesností vylíčeno v beletrii. Dovolte mi představit žraloka, který jedl velryby – skutečné velryby!“
Mnoho dnešních velkých makropredátorských žraloků je známých tím, že pojídají mořské savce, zejména náš starý známý velký bílý žralok. Na některých z nejikoničtějších fotografií tohoto žraloka je vidět, jak se vrhá z vody se svou poslední obětí mezi čelistmi (obr. 1). Mladí žraloci bílí však nejedí tuleně hned od začátku. Místo toho začínají žrát ryby a menší žraloky . Pokud se znovu podíváme na útvar Gatun v Panamě, najdeme spoustu fosilních žraloků, rejnoků a otolitů (struktur z vnitřních uší ryb), stejně jako množství bezobratlých, jako jsou měkkýši .
Něco však nenajdeme, jsou fosilní mořští savci. To zřejmě naznačuje, že mláďata megalodonů se živila také rybami a menšími žraloky, místo aby se hned vrhla na mořské savce. Jedná se o něco, čemu se říká ontogenetický posun v potravě, což znamená, že žralok v průběhu růstu mění preferovaný zdroj potravy. Díky konzumaci většího množství potravy, a to pravděpodobně ve velkém množství, jakmile dosáhl své dospělé velikosti, byl megalodon schopen udržet metabolické nároky spojené s jeho velikostí. Toho mohl dosáhnout nejen pojídáním velryb, ale prakticky čehokoli, na co narazil.

Vzhledem k tomu, kolik potravy musel Megalodon sníst, aby ospravedlnil svou obrovskou velikost, byl téměř jistě oportunistickým žroutem. Fosilní záznamy jsou nám v tomto případě nejlepším vodítkem. Je to dáno přítomností stopových zkamenělin, což jsou geologické záznamy biologické činnosti. Ty mohou sahat od dinosauřích stop až po škrábance na kostech, které naznačují, že zvíře, jemuž kost patřila, něco pokousalo. A megalodon po sobě zanechal několik takových stopových zkamenělin, které poskytují klíčové důkazy o jeho zabití.
Bylo nalezeno několik zkamenělin patřících různým kytovcům s pozoruhodnými stopami po kousnutí. Jeden dobře citovaný příklad pochází z formace Yorktown v Marylandu, kde několik velrybích kostí mělo stopy po kousnutí, které byly zjevně vytvořeny velkými zubatými zuby . Ve skutečnosti se někdy nacházejí zuby megalodona přímo spojené s těmito kostmi .
Žraloci během svého života tisíckrát ztrácejí a vyměňují zuby, a proto žraloci často ztrácejí zuby při násilném zabíjení. To je hlavní důvod, proč jsou žraločí zuby tak častými zkamenělinami. Přítomnost žraločích zubů přímo u poškrábané velrybí kosti skutečně naznačuje, že docházelo ke krmení. Ale něco, co by paleontologové měli při nálezu těchto fosilií vždy zvážit, je: jednalo se skutečně o predátora? Nebo to snad bylo mrchožroutství?
Mrchožroutství je běžné potravní chování u moderních makropredátorských žraloků, jako jsou velcí bílí žraloci (obr. 2) a žraloci tygří . Když tito žraloci narazí na mrtvé velryby plovoucí na hladině, je to pro ně švédský stůl se vším všudy. Kritické je, že mrchožroutství nevyžaduje žádný energetický výdej, který by mohl být promarněn při neúspěšném pokusu o predaci. Vzhledem k tomu, že megalodon potřeboval být schopen udržet si své obrovské rozměry, nedávalo by příliš smysl nechat si ujít jídlo zdarma, pokud by narazil na mrtvou velrybu. Zprostředkovaně podle toho, jak se velcí moderní žraloci živí, by si tedy většina z nás byla ochotna vsadit na to, že Megalodon pravděpodobně mrchožrouty skutečně vyhledával, když měl příležitost.

Podle toho existuje jedna obzvláště pozoruhodná fosilie, která Megalodona usvědčuje z pokusu o predaci. A co je zásadní, víme to především proto, že jeho pokus selhal. V článku publikovaném v roce 2010 byla popsána částečná fosilie žebra pliocénu patřící kytovci z formace Yorktown v Severní Karolíně (ano, dvě formace Yorktown). Tato fosilie rovněž vykazovala poškození připomínající stopy po kousnutí, ale zejména se zdálo, že tyto stopy pokrývá tkaná kost. Autoři to interpretovali jako známku infekce, ke které došlo v době, kdy se zvíře zotavovalo z kostního traumatu způsobeného těmito stopami po kousnutí. Tyto stopy dobře odpovídaly zubatému skusu, což naznačuje, že útočník měl zuby zubaté – stejně jako megalodon. Studie proto naznačuje, že megalodon nebo jiný velký žralok zaútočil na zvíře, z něhož fosilie pochází, ale oběť pokus přežila jen proto, aby o šest týdnů později zemřela na infekci způsobenou zraněním .
Ještě nedávno byly v peruánské formaci Pisco nalezeny vůbec první fosilní stopy po kousnutí na jižní polokouli připisované megalodonovi . Možná to tak mělo být, protože se jedná o stejnou lokalitu, kde Gordon Hubbell našel svou výjimečnou fosilii karcharodona (o níž pojednává 1. čtvrtletí). Zde nalezený vykousnutý materiál obsahoval zbytky lebky a žeber, což naznačuje, že žralok svůj cíl vykousl z boku. Nejvýznamnější je, že pozůstatky byly poprvé identifikovány jako jednotlivý druh – tímto druhem byla Piscobalaena nana, malá velryba. Stejně jako u ostatních stopových fosilií se stopy po kousnutí dobře shodovaly s velkými zuby se zoubky – což je dobrý důvod pro obvinění megalodona z útoku (obr. 3). Ukázalo se tedy, že Megalodon byl schopen zaměřit se i na menší velryby (ať už šlo o dravce, nebo mrchožrouty), což je velmi v souladu s náznaky, že se pravděpodobně zaměřil i na delfíny, mořské želvy a další menší kořist .

Ačkoli je strategie predace z fosilií těžko interpretovatelná, moderní žraloci nám mohou poskytnout představu o tom, jak megalodon lovil. Síla jeho skusu by byla mimořádně silná pro poškození velrybích kostí. Počítačový model lebky žraloka bílého navrhl, že jeho síla skusu přesahovala asi 1,8 tuny, což mu dává jednu z nejsilnějších sil skusu ze všech žijících zvířat . Tato studie následně naznačuje, že síla Megalodonova skusu byla ještě větší a odhaduje se na mimořádných 108 514-182 201 N.
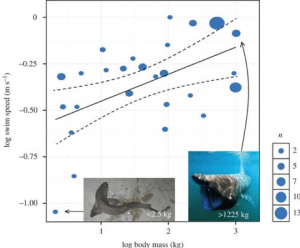
Vědci se také pokusili vypočítat Megalodonovu rychlost plavání, což nám může pomoci naznačit, jakým způsobem lovil. Žraloci mohou mít nárazovou rychlost, což je rychlý výtrysk rychlého plavání, který se obvykle používá k uchopení kořisti. Jejich obvyklou rychlostí při běžném plavání je však to, čemu říkáme trvalá rychlost plavání. Ta se u jednotlivých druhů liší, ale jedna studie vedená Davidem Jacobym zkombinovala údaje o 26 druzích z 64 předchozích studií a vytvořila model, který by předpověděl škálovatelný vztah mezi rychlostí trvalého plavání a metabolismem a tělesnou hmotností žraloka. Jejich model zahrnující metabolismus odhalil škálovací exponent 0,173, shrnutý do následující rovnice:
Rychlost plavání (ms ¹) ∝ Hmotnost (kg)^0,173
Když však korigovali data o fylogenezi, zjistili, že minimální rychlost plavání se škáluje s tělesnou hmotností s o něco nižším exponentem 0,15. V tomto případě se jednalo o model, který byl vytvořen na základě údajů o metabolismu. Výše uvedená rovnice navíc předpokládá konstantu 1, aby odpovídala proporcím, ale údaje ukázaly, že tato konstanta je ve skutečnosti 0,266. Sestavili proto výkonovou rovnici:
Plavecká rychlost (ms ¹) = 0,266 Hmotnost (kg) ^0,15
Z toho jsou schopni vypočítat trvalou rychlost plavání megalodona v závislosti na jeho hmotnosti. Například Megalodon o hmotnosti ~48 000 kg vypočtený v Gottfriedově práci jako hmotnost 15,9 m dlouhého žraloka by ukázal, že žralok plave nepřetržitě rychlostí 1,34 m/s – zhruba 4,8 km/hod . Pokud bychom tento model aplikovali na 52 000 kg vážícího megalodona, zjistili bychom, že jeho trvalá rychlost plavání je 1,36 m/s, tedy 4,9 km/hod, zatímco pokud bychom šli opravdu do extrému a simulovali 100 000 kg vážícího žraloka, výsledkem tohoto modelu by byla rychlost plavání 1,50 m/s, tedy 5,4 km/hod. Tento model se ukázal jako velmi efektivní nejen při výpočtu rychlosti plavání Megalodona, ale také při poskytnutí modelu pro více druhů žraloků (obr. 4) .
Je to však rychlost výbuchu, kterou žraloci využívají k rychlému vytrhnutí kořisti. Žraloci bílí toho využívají při svém „breaking behavior“, kdy se vymrští z vody tak, že se na kořist vrhnou zespodu, aby ji překvapili (obr. 1) . Mohou to dělat, protože jsou dobře maskovaní díky protisvětlu. Protože tedy Megalodon byl pravděpodobně také protistínovaný a měl podobnou ekologii a stavbu těla jako žraloci bílí , nabízí se otázka: Byl Megalodon schopen také breachingu?
Možná. Rychlost výbuchu žraloka bílého je poháněna fyziologickou adaptací zvanou mezotermie . To je něco, co se nedávno předpokládalo, že měl i Megalodon (bude podrobně rozebráno v 5. otázce). Mezotermní ryby mají obecně vyšší trvalou i nárazovou rychlost plavání než ektotermní ryby , a toho bylo využito k domněnce, že Megalodon mohl mít nárazovou rychlost až 37,15 km/hod .
Tato neuvěřitelně vysoká rychlost pro tak velkého žraloka může být dostatečná k tomu, aby byl schopen prorazit. Kromě toho byl pozorován i žralok obrovský (Cetorhinus maximus), druhý největší žralok současnosti, který také proráží . Možná, že obří megalodon byl skutečně schopen vyskočit z vody, když se zmocňoval své poslední potravy. To prostě zatím nevíme, ale můj osobní a (do jisté míry) odborný názor je, že to dost možná bylo možné.
Megalodon byl vrcholovým predátorem, který neměl na výběr z tak širokého jídelníčku, a neměl žádné vlastní známé predátory. To znamená dvě důležité věci. Zaprvé měl pravděpodobně silnou konkurenci v boji o kořist, což mohlo nakonec přispět k jeho vyhynutí. Za druhé, jako vrcholový predátor by byl megalodon pro svůj ekosystém nesmírně důležitý, protože by udržoval populaci své kořisti pod kontrolou, a tím by zachoval zdravý ekosystém.
Proto jsou dnešní žraloci pro mořské prostředí tak důležití. Ztráta vrcholového dravého žraloka vyvolává nevratné kaskádovité účinky, které mění společenství, jež po sobě zanechává . Naše mořské ekosystémy jako takové jsou takové, jaké jsou, částečně proto, že Megalodon již není jejich součástí. A obrovské velryby současných oceánů si díky tomu mohou oddechnout.
- Estrada JA, Rice AN, Natanson LJ & Skomal GB 2006. Využití izotopové analýzy obratlů při rekonstrukci ontogenetické potravní ekologie u žraloků bílých. Ecology 87, 829-834.
- Pimiento C, Ehret DJ, MacFadden BJ & Hubbell G 2010. Starobylá oblast výchovy vyhynulého obřího žraloka Megalodon z miocénu v Panamě. PLoS One 5, e10552.
- Pimiento C, González-Barba G, Ehret DJ, Hendy AJ, MacFadden BJ & Jaramillo C 2013. Žraloci a rejnoci (Chondrichthyes, Elasmobranchii) z pozdně miocenního útvaru Gatun v Panamě. J. Paleontol. 87, 755-774.
- Purdy RW 1996. Paleoecology of fossil white sharks [Paleoekologie fosilních bílých žraloků]. In: Vědecké studie o žralocích a mořských žralocích: Great White Sharks: the biology of Carcharodon carcharias (eds. Klimley AP & Ainley DG), 67-78, Academic Press, San Diego.
- Aguilera OA, García L & Cozzuol MA 2008. Obří zubatí bílí žraloci a trofické interakce kytovců z pliocenní karibské formace Paraguaná. Paläontol. Z. 82, 204-208.
- Fallows C, Gallagher A. & Hammerschlag N 2013. Mrchožroutství žraloků bílých (Carcharodon carcharias) na velrybách a jeho potenciální úloha při dalším utváření ekologie vrcholového predátora. PLoS One, 8, e60797.
- Dudley SFJ, Anderson-Reade MD, Thompson GS & McMullen PB 2000. Concurrent scavenging of a whale carcass by great white sharks, Carcharodon carcharias, and tiger sharks, Galeocerdo cuvier. Fish. Bull. 98, 646-649.
- Kallal RJ, Godfrey SJ & Ortner DJ 2010. Kostní reakce na žebru pliocenního kytovce ukazují na krátkodobé přežití predační události. Int. J. Osteoarchaeol. 22, 253-260.
- Collareta A, Lambert O, Landini W, Di Celma C, Malinverno E, Varas-Malca R, Urbina M & Bianucci G 2017. Zaměřoval se obří vyhynulý žralok Carcharocles megalodon na malou kořist? Stopy po kousnutí na pozůstatcích mořských savců z pozdního miocénu v Peru. Palaeogeogr. Palaeoclimatol. Palaeoecol. 469, 84-91.
- Wroe S, Huber DR, Lowry M, McHenry C, Moreno K, Clausen P, Ferrara TL, Cunningham E, Dean MN & Summers AP 2008. Three-dimensional computer analysis of white shark jaw mechanics: how hard can a great white bite? J. Zool. 276, 336-342.
- Jacoby DM, Siriwat P, Freeman R & Carbone C 2015. Je škálování rychlosti plavání u žraloků podmíněno metabolismem? Biol. Lett. 11, 20150781.
- Gottfried MD, Compagno LJV & Bowman SC 1996. Velikost a anatomie kostry obřího „megatunového“ žraloka Carcharodon megalodon. In: Great White Sharks: the biology of Carcharodon carcharias (eds. Klimley AP & Ainley DG), 55-66, Academic Press, San Diego.
- Jacoby DM, Siriwat P, Freeman R & Carbone C 2016. Correction to „Is the scaling of swim speed in sharks driven by metabolism?“ (Je škálování rychlosti plavání u žraloků podmíněno metabolismem?). Biol. Lett. 12, 20160775.
- Watanabe YY, Payne NL, Semmens JM, Fox A & Huveneers C 2019. Plavecké strategie a energetika endotermních žraloků bílých při hledání potravy. J. Exp. Biol. 222, jeb185603.
- Cooper JA, Pimiento C, Ferrón HG & Benton MJ (v tisku). Tělesné rozměry vyhynulého obřího žraloka Otodus megalodon: A 2D reconstruction. Sci. Rep.
- Watanabe YY, Goldman KJ, Caselle JE, Chapman DD & Papastamatiou YP 2015. Srovnávací analýzy dat ze sledování živočichů odhalují ekologický význam endotermie u ryb. Proc. Nat. Acad. Sci. 112, 6104-6109.
- Ferrón HG 2017. Regionální endotermie jako spouštěč gigantismu u některých vyhynulých makropredátorských žraloků. PLoS One, 12, e0185185.
- Johnston EM, Halsey LG, Payne NL, Kock AA, Iosilevskii G, Whelan B & Houghton JD 2018. Latentní síla žraloků vyhřívajících se odhalená při výjimečných událostech breaking. Biol. Lett. 14, 20180537.
- Myers RA, Baum JK, Shepherd TD, Powers SP & Peterson CH 2007. Kaskádové účinky úbytku vrcholových dravých žraloků z pobřežního oceánu. Science, 315, 1846-1850.
Redakce: Rhys Charles
.
Napsat komentář