Superior olivary nucleus
On 12 listopadu, 2021 by admin| Mozog: Horní olivový komplex | ||
|---|---|---|
 |
||
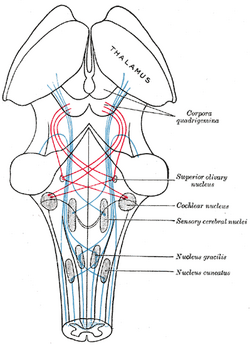
| Schéma znázorňující průběh vláken lemu; mediální lemniscus modře, laterální červeně. (Horní olivové jádro je označeno vpravo uprostřed.) | ||
| ] | ||
| Latinské | jádro olivaris superior | |
| Grayovo | předmět č. 187 787 | |
| část | ||
| Složky | ||
| Arterie | ||
| Vein | ||
| BrainInfo/UW | {{{BrainInfoType}}}-565 | |
| MeSH | ||
Vrchní olivový komplex (neboli SOC nebo vrchní oliva) je malá hmota šedé látky nacházející se na dorzálním povrchu laterální části lichoběžníkového těla. U člověka je malý, ale u některých zvířat dobře vyvinutý, vykazuje podobnou strukturu jako dolní olivové jádro a nachází se bezprostředně nad ním. Některá vlákna trapézového tělesa končí arborizací kolem buněk tohoto jádra, zatímco jiná z těchto buněk vycházejí.
Fyziologie
Vrchní olivové jádro hraje při slyšení řadu rolí. Mediální horní oliva (MSO) je specializované jádro, o němž se předpokládá, že měří časový rozdíl příchodu zvuků mezi ušima (interaurální časový rozdíl neboli ITD). ITD je hlavním vodítkem pro určení azimutu nízkofrekvenčních zvuků, tj. jejich lokalizace v azimutální rovině – jejich stupně vlevo nebo vpravo.
Předpokládá se, že laterální horní oliva (LSO) se podílí na měření rozdílu hladin zvuků mezi ušima (interaurální rozdíl hladin neboli ILD). ILD je druhým hlavním vodítkem při určování azimutu vysokofrekvenčních zvuků.
Vztah ke sluchovému systému
Vrchní olivový komplex se obecně nachází v ponsu, ale u člověka se táhne od rostrální dřeně až do středního ponsu a přijímá projekce převážně z anteroventrálního kochleárního jádra přes ventrální akustickou strunu, ačkoli posteroventrální jádro se tam promítá také přes střední akustickou strunu. V tomto místě dochází k prvním významným binaurálním interakcím.
Primární jádra
Horní olivový komplex se dělí na tři primární jádra, MSO, LSO a mediální jádro trapézového tělesa, a několik menších perioliválních jader. Tato tři jádra jsou nejlépe prozkoumaná, a proto i nejlépe pochopená. Obvykle jsou považována za jádra tvořící vzestupnou azimutální lokalizační dráhu.
Mediální horní oliva (MSO)
Předpokládá se, že mediální horní oliva pomáhá lokalizovat azimut zvuku, tj. úhel vlevo nebo vpravo, kde se nachází zdroj zvuku. Člověka může na první pohled napadnout, že toto jádro zahrnuje vertikální informace, ale není tomu tak; informace zpracovávané v MSO se omezují na horizontální rovinu. Fusiformní buňky se nepromítají do ničeho v úrovni ponsu a vstupují do hry až v dolním koliculu. Jsou zde přítomny pouze horizontální údaje, které však pocházejí ze dvou různých ušních zdrojů, což napomáhá lokalizaci zvuku na ose azimutu. Způsob, jakým to horní oliva dělá, je měření časových rozdílů mezi dvěma ušními signály zaznamenávajícími stejný podnět. Cesta kolem hlavy trvá přibližně 700 μs a předpokládá se, že mediální horní oliva je schopna tuto dobu detekovat. Ve skutečnosti bylo pozorováno, že lidé jsou schopni detekovat interaurální rozdíly až do 10 mikrosekund. Jádro je tonotopicky uspořádané, ale azimutální projekce receptivního pole je „s největší pravděpodobností komplexní, nelineární mapa“.
Projekce mediální horní olivy končí hustě v ipsilaterálním centrálním jádru dolního koliculu. Většina těchto axonů je považována za „kulatý tvar“ neboli typ R. Tyto R axony jsou většinou glutamátergní, obsahují kulaté synaptické vezikuly a tvoří asymetrické synaptické spoje.
- Jedná se o největší z jader a u člověka obsahuje přibližně 15 500 neuronů .
- Každé MSO dostává nízkofrekvenční bilaterální vstupy z pravého a levého AVCN.
- Výstup je do ipsilaterálního laterálního lemnisku a nakonec do dolního koliculu.
- MSO lépe reaguje na binaurální podněty.
- Její hlavní funkce spočívá v detekci rozdílů v době příchodu zvuku k oběma uším, což je součástí lokalizačního procesu (ITD).
Laterální horní oliva (LSO)
Tato oliva má podobné funkce jako mediální horní oliva, ale k lokalizaci zdroje zvuku využívá intenzitu. Je to část mozkového kmene, která označuje hlasitější zvuk z levého ucha jako zvuk z levé strany. Laterální oliva přijímá vstupní informace z obou kochleárních jader, z ipsilaterálních kulovitých buněk AVCN a kontralaterálních kulovitých buněk AVCN prostřednictvím ipsilaterálního MNTB. Hlavní buňky MNTB jsou glycinergní, poskytují zdroj inhibice pro LSO a tvoří základ citlivosti ILD. Projekce z obou kochleárních jader jsou primárně vysokofrekvenční a tyto frekvence jsou následně zastoupeny většinou neuronů LSO (>2/3 v pásmu 2-3 kHz u kočky). Další vstupy pocházejí z ipsilaterálního LNTB (glycinergní, viz níže), které poskytují inhibiční informace z ipsilaterálního kochleárního jádra. Další pravděpodobně inhibiční vstup pochází z ipsilaterálních nesférických buněk AVCN. Tyto buňky jsou buď kulovité keříčkovité, nebo multipolární (hvězdicovité). Každý z těchto dvou vstupů by mohl být základem ipsilaterální inhibice pozorované v mapách odpovědí lemujících primární excitaci a zostřujících frekvenční ladění jednotky.
LSO se bilaterálně promítá do centrálního jádra dolního koliculu (ICC). Ipsilaterální projekce jsou primárně inhibiční (glycinergní) a kontralaterální excitační. Mezi další projekční cíle patří dorzální a ventrální jádro laterálního lemnisku (DNLL & VNLL). GABAergní projekce z DNLL tvoří primární zdroj GABA ve sluchovém mozkovém kmeni a promítají se bilaterálně do ICC a do kontralaterálního DNLL. Tato sbíhající se excitační a inhibiční spojení mohou působit na snížení závislosti citlivosti ILD v ICC na úrovni ve srovnání s LSO.
Další projekce tvoří laterální olivokochleární svazek (LOC), který inervuje vnitřní vláskové buňky kochleárního systému. Předpokládá se, že tyto projekce mají dlouhou časovou konstantu a působí na normalizaci hladiny zvuku detekované každým uchem s cílem pomoci při lokalizaci zvuku. Existují značné druhové rozdíly: U hlodavců jsou projekční neurony LOC rozmístěny uvnitř LSO, u predátorů (např. kočky) LSO obklopují.
Mediální jádro trapézového tělesa (MNTB)
- MNTB je nejmenší částí SOC; přítomnost MNTB v lidském mozkovém kmeni je sporná.
- Každý neuron přijímá pouze vysokofrekvenční vstup prostřednictvím kalichu held z kontralaterálního AVCN a jeho výstup se promítá do ipsilaterálního LSO.
- Nalézáme dva typy odpovědí: „typ chopper“ podobný vřetenitým buňkám v AVCN a primární typ, který je podobný buňkám Bushy v AVCN.
Periolivální jádra
SOC se skládá ze šesti až devíti perioliválních jader, v závislosti na citovaném badateli, obvykle pojmenovaných podle jejich umístění vzhledem k primárním jádrům. Tato jádra obklopují každé z primárních jader a podílejí se na vzestupném i sestupném sluchovém systému. Tato jádra také tvoří zdroj olivokochleárního svazku, který inervuje hlemýžď. U morčete jsou vzestupné projekce do dolních koliculů převážně ipsilaterální (>80 %), přičemž největší jednotlivý zdroj pochází ze SPON. Také ventrální jádra (RPO, VMPO, AVPO, & VNTB) jsou téměř zcela ipsilaterální, zatímco zbývající jádra se promítají bilaterálně.
| Název | Kočka | Morče | Krysa | Myš | ||
|---|---|---|---|---|---|---|
| LSO | X | X | X | X | X | |
| MSO | X | X | X | X | ||
| MNTB | X | X | X | X | X | |
| LNTB | X | X | „LVPO“ | X | ||
| ALPO | X | X | ||||
| PVPO | X | X | ||||
| PPO | X | X | „CPO“ | |||
| VLPO | X | |||||
| DPO | X | X | X | |||
| DLPO | X | X | ||||
| VTB | X | X | „MVPO“ | X | ||
| AVPO | X | |||||
| VMPO | X | X | ||||
| RPO | X | X | ||||
| SPN | „DMPO“ | X | X | X |
Ventrální jádro trapézového tělesa (VNTB)
- VNTB je malé jádro umístěné laterálně od MNTB, a ventrálně od MSO.
- Toto jádro se skládá z heterogenní populace buněk, promítá se do mnoha sluchových jader a tvoří mediální olivokochleární svazek (MOC), který inervuje vnější vláskové buňky kochleárního systému . Tyto buňky obsahují elektromotorická vlákna a v hlemýždi fungují jako mechanické zesilovače/atenuátory.
- Jádro se promítá do obou IC, přičemž žádné buňky se nepromítají bilaterálně.
Laterální jádro trapézového tělesa (LNTB)
- Nachází se ventrálně od LSO
- AVCN kulovité buňky s keříčkovitým uspořádáním promítají kolaterály bilaterálně a kulovité buňky s keříčkovitým uspořádáním promítají kolaterály ipsilaterálně k neuronům LNTB.
- Buňky jsou imunoreaktivní pro glycin a jsou retrográdně označeny po injekci tritiovaného glycinu do LSO
- Jádro se promítá jak do IC, přičemž několik buněk se promítá bilaterálně, tak i do ipsilaterální LSO.
- Velké multipolární buňky se promítají do kochleárního jádra, ale ne do IC, jak u kočky, tak u morčete.
- Vstupy jsou často přes koncové bulby held, což vytváří velmi rychlý přenos signálu.
Horní periolivární jádro (SPON) (dorzomediální periolivární jádro (DMPO)).
- Nachází se přímo dorzálně od MNTB
- U potkana je SPON homogenní GABAergní jádro. Tyto tonotopicky uspořádané neurony přijímají excitační vstupy z osmičkových a multipolárních buněk v kontralaterálním ventrálním kochleárním jádře, glycinergní (inhibiční) vstup z ipsilaterálního MNTB, neznámý GABAergní (inhibiční) vstup a promítají se do ipsilaterálního ICC. Většina neuronů reaguje pouze při posunu podnětu, může fázově blokovat AM podněty až do 200 Hz a může tvořit základ selektivity trvání ICC. Pozoruhodné je, že neurony SPON nedostávají sestupné vstupy z IC a nepromítají se do hlemýždě nebo kochleárního jádra jako mnohá periolivární jádra.
- U morčete se kulaté až oválné multipolární buňky promítají do obou IC, přičemž mnoho buněk se promítá bilaterálně. Podlouhlejší buňky, které se promítají do kochleárního jádra, se do ICC nepromítají. Zdá se, že existují dvě populace buněk, jedna, která se promítá ipsilaterálně, a druhá, která se promítá bilaterálně.
- Většina informací pocházela ze SPON hlodavců, vzhledem k nápadné velikosti jádra u těchto druhů, přičemž u kočičího DMPO bylo provedeno jen velmi málo studií, z nichž žádná nebyla rozsáhlá.
Dorzální periolivální jádro (DPO)
- Umístěno dorzálně a mediálně od LSO
- Obsahuje jednotky EE (buzené oběma ušima) i E0 (buzené pouze kontralaterálním uchem).
- Neurony jsou tonotopicky uspořádané a vysokofrekvenční.
- Může patřit k jednomu jádru spolu s DLPO
- Jádro promítá do obou IC, přičemž žádné buňky nepromítají bilaterálně.
Dorsolaterální periolaterální jádro (DLPO)
- Nachází se dorzálně a laterálně od LSO
- Obsahuje jednotky EE (buzené oběma ušima) i E0 (buzené pouze kontralaterálním uchem).
- Neurony jsou tonotopicky uspořádané a nízkofrekvenční.
- Může patřit k jednomu jádru spolu s DPO
- Jádro promítá do obou IC, jen málo buněk promítá bilaterálně.
Ventrolaterální periolivální jádro (VLPO)
- Nachází se ventrálně a uvnitř ventrálního hilu LSO
- Obsahuje jednotky EI (buzené kontralaterálním a inhibované ipsilaterálním uchem) i E0 (buzené pouze kontralaterálním uchem).
- Neurony jsou tonotopicky uspořádané a mají vysokou frekvenci.
Anterolaterální periolivální jádro (ALPO)
- Jádro promítá do obou IC, přičemž žádné buňky nepromítá bilaterálně.
- Velké multipolární buňky se promítají do kochleárního jádra, ale ne do IC, a to jak u kočky, tak u morčete.
Ventromediální periolivární jádro (VMPO)
- Nachází se mezi MSO a MNTB.
- Oboustranně vysílá projekce do ICC.
- Jádro promítá do obou IC, přičemž žádné buňky nevysílají projekce bilaterálně.
Rostrální periolaterální jádro (RPO) (Přední periolaterální jádro (APO)).
- Nachází se mezi rostrálním pólem MSO a VNLL
Kaudální periolivární jádro (CPO) (zadní periolivární jádro (PPO)).
- Nachází se mezi kaudálním pólem MSO a obličejovým jádrem (7N)
Posteroventrální periolivární jádro (PVPO)
- Jádro vystupuje do obou IC, přičemž žádné buňky nevystupují bilaterálně.
Patofyziologie
Při pitvě 21leté ženy s autismem, epilepsií a mentální retardací byla zjištěna téměř úplná absence horní olivy.
Viz také
- Olivary body
- Kulesza RJ, Cytoarchitecture of the human superior olivary complex: Medial and lateral superior olive. Hearing Research 225(2007) 80-90
- 2.0 2.1 Oliver DL, et al. Axonal projections from the lateral and medial superior olive to the inferior colliculus of the cat: a study using electron microscopic autoradiography. J Comp Neurol. 1995 Sep 11;360(1):17-32 Citovat chybu: Cajal, S. R. Y. and L. Azoulay (1909): Invalid
<ref>tag; name „R4“ defined multiple times with different content - . Histologie du système nerveux de l’homme et des vertébrés. Paris, Maloine.
- 4.0 4.1 Kandel, et al Principles of Neuroscience. Čtvrté vydání. s. 591-624. Copyright 2000, by McGraw-Hill Co. Citovat chybu: Kulesza, R. J., Jr.: Invalid
<ref>tag; name „R2“ defined multiple times with different content - Kulesza, R. J., Jr. (2007). „Cytoarchitektura lidského horního olivového komplexu: mediální a laterální horní oliva“. Hear Res 225(1-2): 80-90.
- Tsuchitani, C. a J. C. Boudreau (1967). „Encoding of stimulus frequency and intensity by cat superior olive S-segment cells“ (Kódování frekvence a intenzity podnětů buňkami S-segmentu horní olivy kočky). J Acoust Soc Am 42(4): 794-805.
- 8.0 8.1 8.2 Chyba při citování: Invalid
<ref>tag; name „R10“ defined multiple times with different content - Wu, S. H. and J. B. Kelly (1994). „Fyziologický důkaz ipsilaterální inhibice v laterální horní olivě: synaptické odpovědi v plátku myšího mozku“. Hear Res 73(1): 57-64.
- Brownell, W. E., P. B. Manis a další (1979). „Ipsilaterální inhibiční odpovědi v laterální horní olivě kočky“. Brain Res 177(1): 189-93.
- Darrow, K. N., S. F. Maison a další (2006). „Cochlear efferent feedback balances interaural sensitivity“. Nat Neurosci 9(12): 1474-6.
- Warr, W. B. a J. J. Guinan, Jr. (1979). „Eferentní inervace cortiho orgánu: dva oddělené systémy“. Brain Res 173(1): 152-5.
- 13.0 13.1 Schofield, B. R. a N. B. Cant (1991). „Organizace horního olivového komplexu u morčete. I. Cytoarchitektura, histochemie cytochromoxidázy a morfologie dendritů“. J Comp Neurol 314(4): 645-70. Citovat chybu: Invalid
<ref>tag; name „R7“ defined multiple times with different content - 14.0 14.1 14.2 14.3 14.4 14.5 14.6 14.7 14.8 Illing, R. B., K. S. Kraus, et al. (2000). „Plasticita horního olivového komplexu“. Microsc Res Tech 51(4): 364-81. Citovat chybu: Cite error: Invalid
<ref>tag; name „R5“ defined multiple times with different content Chyba citace: Cite error: Invalid<ref>tag; name „R5“ defined multiple times with different content Chyba citování: Invalid<ref>tag; name „R5“ defined multiple times with different content Chyba citace: Invalid<ref>tag; name „R5“ defined multiple times with different content Chyba citace: Invalid<ref>tag; name „R5“ defined multiple times with different content Chyba citace: Invalid<ref>tag; name „R5“ defined multiple times with different content Chyba citace: Invalid<ref>tag; name „R5“ defined multiple times with different content Chyba citace: Warr, W. B. and J. E. Beck (1996): Invalid<ref>tag; name „R5“ defined multiple times with different content - . „Vícenásobné projekce z ventrálního jádra trapézového tělíska u potkana“. Hear Res 93(1-2): 83-101.
- 16.00 16.01 16.02 16.03 16.04 16.05 16.06 16.07 16.08 16.09 16.10 Schofield, B. R. a N. B. Cant (1992). „Organization of the superior olivary complex in the guinea pig: II. Patterns of projection from the periolivary nuclei to the inferior colliculus“. J Comp Neurol 317(4): 438-55. Cite error: Cite error: Invalid
<ref>tag; name „R6“ defined multiple times with different content Chyba citace: Invalid<ref>tag; name „R6“ defined multiple times with different content Chyba citace: Invalid<ref>tag; name „R6“ defined multiple times with different content Chyba citace: Invalid<ref>tag; name „R6“ defined multiple times with different content Chyba citace: Invalid<ref>tag; name „R6“ defined multiple times with different content Chyba citace: Invalid<ref>tag; name „R6“ defined multiple times with different content Chyba citace: Invalid<ref>tag; name „R6“ defined multiple times with different content Chyba citace: Invalid<ref>tag; name „R6“ defined multiple times with different content Chyba citace: Invalid<ref>tag; name „R6“ defined multiple times with different content Cite error: Invalid<ref>tag; name „R6“ defined multiple times with different content Cite error: (1993): Invalid<ref>tag; name „R6“ defined multiple times with different content - Smith, P. H., P. X. Joris, et al. „Projekce fyziologicky charakterizovaných axonů kulovitých křovinatých buněk z kochleárního jádra kočky: důkaz zpožděných linií do mediální horní olivy“. J Comp Neurol 331(2): 245-60.
- Wenthold, R. J., D. Huie a další (1987). „Glycinová imunoreaktivita lokalizovaná v kochleárním jádru a komplexu horní olivy“. Neuroscience 22(3): 897-912.
- 19.0 19.1 Adams, J. C. (1983). „Cytologie perioliválních buněk a organizace jejich projekcí u kočky“. J Comp Neurol 215(3): 275-89. Cite error: Invalid
<ref>tag; name „R8“ defined multiple times with different content - Friauf, E. and J. Ostwald (1988). „Divergentní projekce fyziologicky charakterizovaných neuronů ventrálního kochleárního jádra potkana prokázané intraaxiální injekcí křenové peroxidázy“. Exp Brain Res 73(2): 263-84.
- Kulesza, R. J., Jr. a A. S. Berrebi (2000). „Superior paraolivary nucleus of the rat is a GABAergic nucleus“. J Assoc Res Otolaryngol 1(4): 255-69.
- Kulesza, R. J., Jr, G. A. Spirou a další (2003). „Fyziologické vlastnosti odezvy neuronů v horním paraolivaryngu potkana“. J Neurophysiol 89(4): 2299-312.
- WHITE JS, WARR WB. The dual origins of the olivocochlear bundle in the albino rat [Dvojí původ olivokochleárního svazku u albínského potkana]. J. Comp. Neurol. 219:203-214, 1983.
- Guinan, J. J., Jr., S. S. Guinan a další (1972). „Single auditory units in the superior olivary complex. I. Reakce na zvuky a klasifikace na základě fyziologických vlastností“. Int J Neurosci 4: 101-20.
- Tsuchitani, C. (1977). „Functional organization of lateral cell groups of cat superior olivary complex“. J Neurophysiol 40(2): 296-318.
- Rodier PM, Ingram JL, Tisdale B, Nelson S, Romano J (1996). Embryologický původ autismu: vývojové anomálie motorických jader kraniálních nervů. J Comp Neurol 370 (2): 247-61.
- MedEd at Loyola Neuro/frames/nlBSs/nl27fr.htm
- Obrázek na med.utah.edu (výběr 9b. Pons)
Tento článek byl původně založen na záznamu z veřejně dostupného vydání Grayovy anatomie. Proto mohou být některé informace v něm obsažené zastaralé. Pokud tomu tak je, upravte prosím článek a neváhejte odstranit tuto poznámku, pokud již není relevantní.
|
v-d-e
Mozek: rhombencephalon (zadní mozek) |
||||||
|---|---|---|---|---|---|---|
| Metencephalon/ pons |
|
Dorsal/
(tegmentum) |
Ventrální/
(báze) |
Raphe/retikulární
|
||
| Myelencephalon/ medulla |
|
Dorsální
|
Ventrální
|
Raphe/retikulární
|
||
|
v-d-e
Smyslový systém: Sluchový a vestibulární systém (TA A15.3, GA 10.1029) |
||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Vnější ucho |
Pinna (Helix, Antihelix, Tragus, Antitragus, Incisura anterior auris, Earlobe) – Ušní kanálek – Ušní svaly Eardrum (Umbo, Pars flaccida) |
|||||||||||
| Střední ucho |
|
|||||||||||
| Vnitřní ucho/ (membránový labyrint, kostní labyrint) |
|
|||||||||||
| {| class=“navbox collapsible nowraplinks“ style=“margin: | ||||||||||||
|
—
|
||||||||||||
|
||||||||||||
|}
Tato stránka využívá obsah s licencí Creative Commons z Wikipedie (zobrazit autory).
Napsat komentář