Struktura a funkce bakteriálních buněk
On 10 října, 2021 by adminStruktura a funkce bakteriálních buněk (strana 5)
(Tato kapitola má 10 stran)
© Kenneth Todar, PhD
Buněčná stěna
Buněčné stěny bakterií si zaslouží zvláštní pozornost z několika důvodů:
1. Jsou základní strukturou pro životaschopnost, jak bylo popsáno výše.
2. Jsou složeny z unikátních složek, které se nikde jinde v přírodě nevyskytují.
3. Jsou jedním z nejdůležitějších míst pro útok antibiotik.
4. Poskytují ligandy pro adherenci areceptory pro léčiva nebo viry.
5. Jsou jedním z nejdůležitějších míst pro útok antibiotik. Způsobují příznakyonemocnění u zvířat.
6. Zajišťujíimunologické rozlišenía imunologické rozdíly mezi kmeny bakterií.
Většina prokaryot má tuhou buněčnou stěnu. buněčná stěna je základní struktura, která chrání buněčnýprotoplastpřed mechanickým poškozením a před osmotickým roztržením nebo lýzou.Prokaryota obvykle žijí v relativně zředěném prostředí, takže akumulace rozpuštěných látek uvnitř cytoplazmy prokaryotické buňky výrazně převyšuje celkovou koncentraci ve vnějším prostředí. Tak může být osmotickýtlakna vnitřek plazmatické membrány ekvivalentní10-25atm. Protože membrána je křehká, plastická struktura, musí být omezena vnější stěnou z porézního, pevného materiálu, který má vysokou pevnost v tahu. Takovým materiálem je murein, všudypřítomná složkabakteriálních buněčných stěn.
Murein je jedinečný typ peptidoglykanu, polymeru disacharidů (glykanu) zesíťovaného krátkými řetězciaminokyselin (peptidu). Existuje mnoho typů peptidoglykanu. Všechny bakteriálnípeptidoglykany obsahují kyselinu N-acetylmuramovou, která je určující složkou mureinu. Buněčné stěny archeí mohou být složeny z bílkovin, polysacharidů nebo molekul podobných peptidoglykanu, ale nikdy neobsahují murein. Tato vlastnost odlišuje bakterie od archeí.
U grampozitivních bakterií (těch, které při barvení podle Grama zachovávají fialové barvivo krystalovou violeť) se buněčná stěna skládá z několika vrstev peptidoglykanu. Kolmo k peptidoglykanovým vrstvám probíhá skupina molekul zvaných teichoové kyseliny, které jsou pro grampozitivní buněčnou stěnu jedinečné (obrázek 14). 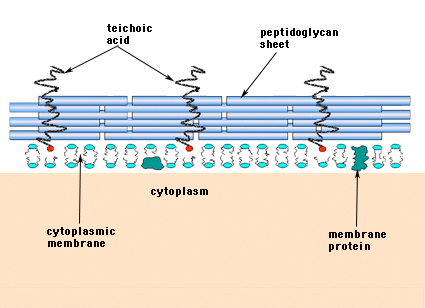
Obrázek 14. Struktura buněčné stěny grampozitivních bakterií. Stěna je relativně silná a skládá se z mnoha vrstev peptidoglykanuproložených teichoovými kyselinami, které probíhají kolmo na listy peptidoglykanu.
U gramnegativních bakterií (které nezachycují krystalovou violeť) se buněčná stěna skládá z jediné vrstvy peptidoglykanůobklopené membránovou strukturou zvanou vnější membrána.Vnější membrána gramnegativních bakterií vždy obsahuje jednosložkový lipopolysacharid (LPS nebo endotoxin), který je pro zvířata toxický. U gramnegativních bakterií je vnější membrána obvykle považována za součást buněčné stěny (obrázek15).

Obrázek15. Struktura gramnegativní buněčné stěny. Stěna je relativnětenká a obsahuje mnohem méně peptidoglykanu než stěna grampozitivní. chybí také teichoové kyseliny. Gramnegativní buněčná stěna se však skládá z vnější membrány, která je mimo peptidoglykanovou vrstvu. Vnější membrána je k peptidoglykanové vrstvě připojena pomocí jedinečné skupiny lipoproteinových molekul.
U grampozitivních bakterií je buněčná stěna silná (15-80 nanometrů) a skládá se z několika vrstev peptidoglykanu. U gramnegativních bakterií je buněčná stěna relativně tenká (10 nanometrů) a skládá se z jediné vrstvy peptidoglykanů obklopené vnější membránou.
Struktura a uspořádání peptidoglykanu u E.coli je reprezentativní pro všechny Enterobacteriaceae i pro mnoho dalších gramnegativních bakterií. Glykanová páteř se skládá ze střídajících se molekul N-acetylglukosaminu (G) a N-acetylmuraminové kyseliny (M) spojených beta 1,4-glykosidovou vazbou. Tříuhlík kyseliny N-acetylmuramové (M) je nahrazen laktyletherovou skupinou odvozenou od pyruvátu. Laktylether spojuje glykanovou páteř s postranním řetězcem peptidu, kterýobsahujeL-alanin, (L-ala), D-glutamát (D-glu), kyselinu diaminopimelovou (DAP) aD-alanin (D-ala). MurNAc je jedinečný pro bakteriální buněčné stěny, stejně jakoD-glu, DAP a D-ala. Podjednotka kyseliny muraminové E. coli je znázorněna naobrázku16 níže.

Obrázek 16. Struktura podjednotky kyseliny muramové peptidoglykanu Escherichia coli. jedná se o typ mureinu, který se nachází u většiny gramnegativních bakterií. Základemglykanu je opakující se polymer dvou aminocukrů, N-acetylglukosaminu (G) a N-acetylmuraminové kyseliny (M). K N-acetylmuramové kyselině je připojen atetrapeptid sestávající z L-ala-D-glu-DAP-D-ala. b. Zkrácená struktura podjednotky kyseliny muramové. c. Blízké tetrapeptidové postranní řetězce mohou být navzájem spojeny interpeptidovou vazbou mezi DAP na jednom řetězci a D-ala na druhém. d. Polymerní forma molekuly.
Vlákna mureiny jsou sestavena vperiplasmě z přibližně 10 podjednotek kyseliny muramové. Poté se vlákna propojí a vytvoří souvislou glykanovou molekulu, která obklopuje buňku. tam, kde to jejich blízkost umožňuje, mohou být tetrapeptidové řetězce, které vystupují z glykanové páteře, zesíťovány interpeptidovou vazbou mezi volnou aminoskupinou na DAP a volnou karboxyskupinou na blízké D-ale.Složení peptidoglykanu na vnější straně plazmatické membrány je zprostředkováno skupinou periplazmatických enzymů, kterými jsou transglykosylázy, transpeptidázy a karboxypeptidázy. Mechanismus účinku penicilinu a příbuzných beta-laktamových antibiotik spočívá v blokování transpeptidázových a karboxypeptidázových enzymů během jejich sestavování mureinové buněčné stěny. Proto se o betalaktamových antibiotikách říká, že „blokují syntézu buněčné stěny“ vbakteriích.
Glykanovou páteř peptidoglykanové molekuly může štěpit enzym zvaný lysozym, který je přítomen vživočišnémséru, tkáních a sekretech a ve fagocytárním lysozomu. Funkcí lysozymu je lyzovat bakteriální buňky jako konstitutivní obrana proti bakteriálním patogenům. Některé grampozitivní bakterie jsou na lyzozym velmi citlivé a enzym je poměrně aktivní již při nízkých koncentracích. lyzozymové sekrety (slzy) lze ředit v poměru 1:40 000 a zachovávají si schopnost lyzovat některé bakteriální buňky. Gramnegativní bakterie jsou vůči útoku lysozymu méně zranitelné, protože jejich peptidoglykan je chráněn vnější membránou. Přesným místem štěpení lysozymu je vazba beta 1,4 mezi kyselinouN-acetylmuramovou (M) a N-acetylglukosaminem (G) , takže podjednotka kyselinymuramové znázorněná na obrázku 16a) je výsledkem působení lysozymu na bakteriální peptidoglykan.
U grampozitivních bakterií existuje mezi peptidoglykany řada různých uspořádání peptidů. Nejlépe prozkoumaná jemureina Staphylococcus aureus zobrazená na obrázku 17 níže. MístoDAP(uE. coli) je diaminokyselina L-lysin (L-lys) a místo interpeptidové vazby (u gramnegativních) je interpeptidový můstek aminokyselin, který spojuje volnou aminoskupinu na lysinu s volnoukarboxyskupinou na D-ale blízkého postranního řetězce tetrapeptidu. Toto uspořádání zřejmě umožňuje častější křížové vazby mezi blízkými postranními řetězci tetrapeptidů. U S. aureus je interpeptidový můstek tvořen peptidem složeným z 5 molekul glycinu (tzv. pentaglycinový můstek). Sestavení interpeptidového můstku u grampozitivního mureinu je inhibováno beta-laktamovými antibiotiky stejným způsobem jako interpeptidová vazba u gramnegativního mureinu. Grampozitivní bakterie jsou citlivější kpenicilinůmnež gramnegativní bakterie, protože peptidoglykan není chráněn vnější membránou a je to hojnější molekula. UGrampozitivníchbakterií se peptidoglykany mohou lišit v aminokyselině na místě DAP neboL-lys v poloze 3 tetrapeptidu a v přesném složení interpeptidového můstku. U grampozitivních bakterií existuje nejméně osm různých typůpeptidoglykanů.
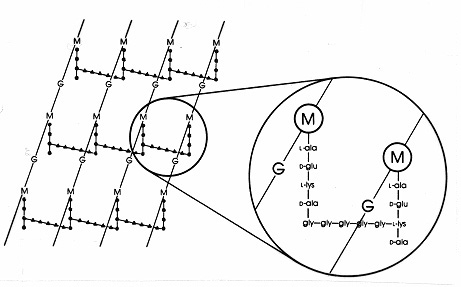
Obrázek 17. Schematické schéma peptidoglykanového listu Staphylococcus aureus. G = N-acetyl-glukosamin;M = N-acetyl-muraminová kyselina; L-ala = L-alanin; D-ala = D-alanin; D-glu = D-glutamová kyselina; L-lys = L-lysin. Jedná se o jeden typ mureinu, který se vyskytuje vGram-pozitivních bakteriích. Ve srovnání s peptidoglykanem E. coli (obr. 7) je v tetrapeptidu místo DAP (kyseliny diaminopimelové) L-lys. volná aminoskupina L-lys je nahrazena glycin-pentapeptidem (gly-gly-gly-gly-gly-), který se pak stává interpeptidovým můstkem tvořícím spojení s karboxylovou skupinou z D-ala v sousedním postranním řetězci tetrapeptidu. Grampozitivní peptidoglykany se liší druh od druhu,především co se týče aminokyselin ve třetí pozici tetrapeptidového řetězce a aminokyselinového složení interpeptidového můstku.
Gramnegativní bakterie mohou ve svých buněčných stěnách obsahovat jednu monomolekulární vrstvu mureinu, zatímco u grampozitivních bakterií se předpokládá, že mají několik vrstev nebo „obalů“ peptidoglykanu. S vrstvami peptidoglykanu u grampozitivních bakterií je úzce spojena skupina molekul zvaných teichoové kyseliny. Teichoové kyseliny jsou lineární polymery polyglycerolu nebo polyribitolu substituované fosfáty a několika aminokyselinami a cukry. Polymery teichoových kyselin jsou občas ukotveny v plazmatické membráně (nazývané lipoteichoová kyselina, LTA), zřejmě směrované ven v pravém úhlu k vrstvám peptidoglykanu. Funkce kyselinyteichoové nejsou známy. Ve volné přírodě jsou nezbytné pro životaschopnostGram-pozitivníchbakterií. Jednou z myšlenek je, že poskytují kanál s pravidelně orientovanými zápornými náboji pro průchod kladně nabitých látek složitou peptidoglykanovou sítí. Další teorií je, že se teichokyseliny nějakým způsobem podílejí na regulaci a sestavování podjednotek muramových kyselin na vnější straně plazmatické membrány. Existují případy,zejména u streptokoků, kdy se teichoové kyseliny podílejí na přilnavosti bakterií k povrchu tkání.
kapitola pokračování
Předchozí strana
Napsat komentář