Frontiers in Psychology
On 28 září, 2021 by adminTělo mozkové (corpus callosum, CC) je primární komisura spojující obě mozkové hemisféry. Proto, i když termín kommissurotomie není nesprávný pro popis takzvaného „rozdělení mozku“ a je striktně správný pro původní chirurgické operace u člověka, při nichž byly všechny komisury přerušeny (Gazzaniga et al., 1967), pozdější operace odřízly pouze CC, čímž se začal používat termín „kalosotomie“, který v těchto případech zůstává jako vhodnější (např. Franz et al., 1996, 2000).
Z evolučního hlediska je CC recentním vývojem pozorovaným pouze u placentálních savců (Katz et al., 1983; Mihrshahi, 2006). Předpokládá se, že funkci, kterou CC zajišťuje, alespoň částečně předcházela přední komisura (AC), pozorovaná u neplacentárních savců, jako jsou vačnatci (Heath a Jones, 1971; Ashwell et al., 1996). Ačkoli jsou v lidském mozku stále přítomny některé zbytkové dráhy AC, její konektivita a struktura zůstává v současné literatuře nedostatečně prozkoumána a řídké nálezy jsou spíše rozporuplné (Lasco et al., 2002; Patel et al., 2010; Choi et al., 2011). Například nálezy založené na zobrazování magnetickou rezonancí (MRI) se rozcházejí v tom, zda má AC podobnou velikost u žen a mužů (Lasco et al., 2002; Patel et al., 2010). V relativně nedávné studii využívající in-vivo techniku difuzního tenzorového zobrazování (DTI), která využívá difúzi molekul vody v axonech, Patel et al. (2010) studovali osm subjektů a úspěšně izolovali vlákna AC u pěti subjektů. Mezi zjištěními byl u jednoho subjektu velký podíl vláken v zadní končetině AC putujících oboustranně do parietálních oblastí, kde většina z nich končila, přičemž menší svazek končil v precentrálním gyru a další svazek vstupoval do temporálního laloku. Takovéto individuální rozdíly byly zjištěny u všech účastníků, přesto většina subjektů vykazovala bilaterální projekce do orbitofrontálního, parietálního a temporálního kortexu, do kterých se KK jednoznačně nepromítá. Dohromady tato zjištění naznačují, že AC má primární i vestigiální projekce do neokortikálních oblastí, kam se CC nepromítá, jak předpokládají i jiní (Di Virgilio et al., 1999; Patel et al., 2010).
Podle našeho názoru AAA do značné míry spoléhá na neokortikální sítě a konektivitu s podkorovými strukturami, jako jsou bazální ganglia a thalamus (mimo jiné, které nejsou v našem návrhu konkrétně diskutovány) (Franz, 2012). Jakýkoli evoluční vývoj zahrnující AC by tedy mohl hrát určitou roli i v modelu AAA. Našli jsme však jen málo literatury, která by se konkrétně zabývala touto možnou souvislostí.
Úloha AC byla studována v úlohách, které pravděpodobně vyžadují formy pozornosti, jako jsou úlohy zahrnující zrakovou pozornost. Je například dobře známo, že populace akallosy byly studovány v souvislosti s úlohami zahrnujícími vizuální integraci (Gazzaniga et al., 1962, 1965; Sperry, 1968; Levy et al., 1972). V nedávné době se Corballis (1995) snažil zabývat vizuální integrací a pozorností ve snaze vysvětlit předchozí rozpory ve výzkumu akallosy. Corballis navrhl dichotomický systém zrakové pozornosti; jedna část (zahrnující formu) je automatická a lokalizovaná v každé hemisféře a druhá (zahrnující pohyb a umístění) je dobrovolná a podkorová. Model AAA může vycházet z tohoto pozorování a řešit nesrovnalosti pozorované v literatuře. Na základě námi navrženého modelu existují na více úrovních pozornosti i jiné cesty než CC, které mohou zprostředkovávat přenos informací mezi hemisférami. Jedna z možností zahrnuje obvody bazálních ganglií, které podle našeho návrhu mají potenciálně rozhodující význam při zprostředkování sdílení pozornosti mezi hemisférami (Franz, 2012). Přerušením CC je přímý přenos pozornosti prostřednictvím korových drah do značné míry zrušen (pokud ovšem AC nehraje záložní roli). Přesto může být integrace zrakových informací založená na pozornosti stále zprostředkována prostřednictvím podkorových drah a/nebo AC. Tímto způsobem by mohl být přítomen dvoudílný systém pozornosti pozorovaný Corballisem (1995) v důsledku zprostředkování vizuální integrace na základě pozornosti prostřednictvím AC (a/nebo bazálních ganglií a zapojených okruhů: Franz, 2012).
Studie zahrnující zrak a pozornost není třeba posuzovat zcela odděleně od systému regulujícího motorickou akci. Na základě našich vlastních zjištění (Franz, 2004; Franz a Packman, 2004) jsou různé formy pozornosti (vizuální a interní-nevizuální) úzce propojeny s motorickou akcí a je logické, že možný klíč k pochopení potenciální role AC v oblasti pozornosti spočívá ve spojení mezi akcí, zrakem a pozorností. V dřívější studii nechal Franz (2004) normální účastníky bimanuálně kreslit kruhy při různých manipulacích s vizuální zpětnou vazbou nebo vnitřní pozorností na každou ruku. Velikost kruhů nakreslených rukou, které se dostávalo určité formy vnitřní pozornosti, se zvětšila podobným způsobem jako účinky, které se projevily při přítomnosti vizuální zpětné vazby této ruky (ve srovnání s absencí pozornosti/vizuální zpětné vazby). Náš model AAA implikuje pozornost jako klíčovou složku v regulaci motorického výstupu (stejně jako výběr toho, jaká motorická akce je plánována), a pokud jsou k dispozici současné důkazy, nemůžeme vyloučit, že AC hraje v této funkci určitou roli. Přímé testování role AC v jakémkoli kognitivním úkolu však není triviální.
Testování, zda se AC podílí na kognitivní funkci, je v neuropsychologickém výzkumu téměř nemožné vzhledem k tomu, že účastníky s lokalizovanou AC není snadné najít (a my o žádném nevíme). Mnohem intuitivnější by proto mohlo být posoudit neurologické rozdíly v normálním AC a korelovat neurologické údaje s údaji o chování. Předloženou traktografickou metodu (objasňující AC) lze poměrně snadno aplikovat na velkých vzorcích normální populace a v ideálním případě ji lze kombinovat s výkony (na základě úkolů prováděných během samostatných testovacích sezení ze sbírky DTI) týchž jedinců pomocí účelově navržených kognitivních úloh. V tomto článku demonstrujeme, že DTI lze provádět a že může přinést použitelné výsledky na malém vzorku osob z neurologicky normální populace. Obecným návrhem by bylo provést podobné metody na větším vzorku a (jak bylo navrženo výše), korelovat zjištění (například proxy míry hustoty AC) s proměnnými v kognitivní úloze.
Na obrázku 1 ukazujeme výsledky sledování DTI na datech získaných od 10 účastnic získaných z veřejného souboru dat DTI na Johns Hopkins University (úplný zdroj viz Poděkování). Průměrný věk byl 22 let (SD = 2,5). Snímky byly zpracovány pomocí FSL Diffusion Toolbox (Smith et al., 2004; Woolrich et al., 2009; Jenkinson et al., 2012) a byly použity postupy použité v předchozích studiích na jiných vláknových drahách (FDT; Behrens et al., 2003a,b, 2007; Johansen-Berg et al., 2004). Stručně řečeno, snímky byly nejprve spojeny do 4d objemů pro každého účastníka a lebky byly rozebrány pomocí nástroje FSL Brain Extraction Tool (BET; Smith, 2002; Jenkinson et al., 2005). Extrahované obrazy byly korigovány na drobné pohyby hlavy a vířivé proudy vzniklé změnami směru pole. Difuzní tenzory byly přizpůsobeny k vytvoření snímků, které zobrazují primární směry difuze a úrovně FA v každém voxelu, aby mohly být použity pro barevné mapy při maskování. Bayesovský odhad parametrů difuze získaných pomocí technik vzorkování s křížícími se vlákny (BEDPOSTX; „X“ znamená křížící se vlákna) byl proveden za účelem vytvoření distribuce velikosti a směru difuze zohledňující dvě křížící se vlákna pro pozdější provedení traktografie napříč difuzními voxely.
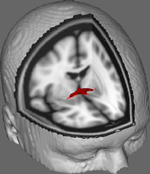
Obrázek 1. Zprůměrované výsledky sledování u 10 kontrolních účastníků překryté na standardní T1 vážený MRI obraz s 2 mm izotropními voxely pro anatomickou referenci.
Zprůměrované výsledky sledování u všech 10 kontrolních účastníků byly poté překryty na standardní T1 vážený MRI obraz s 2 mm izotropními voxely pro anatomickou referenci. Průměrný poměr FA ve skupině byl 0,3146 (SD = 0,0472). Tyto výsledky traktografie ukazují, což je důležité, že strukturu AC lze objasnit, a proto má potenciál pro další studie korelující skóre výkonu v behaviorálních úkolech s hodnotami FA za použití většího vzorku.
Shrnem lze říci, že v současné době existuje řídká literatura a z dostupné literatury žádná konkrétně nespojuje AC s procesy souvisejícími s modelem AAA, na který se zaměřuje náš dřívější cílový článek (Franz, 2012). Pomocí postupu podobného zde navrženému je možné testovat větší vzorky normální populace a korelovat hodnoty FA výsledků DTI (například) s konkrétní závislou proměnnou získanou z účelově vyvinutého úkolu zaměřeného na pozornost k činnosti (jehož postupy jsou v naší laboratoři ve vývoji).
Prohlášení o střetu zájmů
Autoři prohlašují, že výzkum byl prováděn bez jakýchkoli komerčních nebo finančních vztahů, které by mohly být chápány jako potenciální střet zájmů.
Poděkování
Vděčně děkujeme Human Brain Project a National Research Resource Center (USA) a John Hopkins University za umožnění veřejného použití jejich dat DTI. Jsme také vděčni za štědrou grantovou podporu Marsdenova fondu Královské společnosti Nového Zélandu (Elizabeth A. Franz).
Corballis, M. (1995). Vizuální integrace v rozděleném mozku. Neuropsychologia 33, 937-959.
Pubmed Abstract | Pubmed Full Text
Franz, E. A. (2012). Alokace pozornosti při učení cíleně zaměřených činností: rámec kognitivní neurovědy se zaměřením na bazální ganglia. Front. Psychol. 3:535. doi: 10.3389/fpsyg.2012.00535
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Heath, C., and Jones, E. (1971). Interhemisférické dráhy při absenci corpus callosum. An experimental study of commissural connexions in the marsupial phalanger. J. Anat. 109, 253-270.
Pubmed Abstract | Pubmed Full Text
.
Napsat komentář